
5.2: מארגנים - מארגנים אחרים
- Page ID
- 207651

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מארגנים
אחד הדברים המעניינים ביותר בבניית גופים של בעלי חיים הוא המגוון שאנו רואים על פני ובתוך גופים. לדוגמה, למרות שדפוס הגן של הוקס בזבובי פירות ובעכברים דומה מאוד, התוצאות הסופיות (עכבר בוגר או זבוב בוגר) שונות בתכלית. אנו רואים גיוון מסקרן גם בתוך גופים. לדוגמה, קטע בית חזה בזבוב פירות עשוי לבטא גן Hox יחיד על פני מקטע, אך חלקים מקטע זה לובשים צורות רבות ושונות. חלק ניכר מההבחנה הזו נשלטת על ידי מארגנים מקומיים וגנים לשליטה מאסטר. אלה גנים המתבטאים באופן מקומי המתבטאים לעתים קרובות בקטעים רבים בנקודות דומות. בהתאם לגן הוקס המתבטא בקטע זה, הם יפעילו חלקי גוף שונים. לדוגמה, נסתכל בהמשך על גנים לבקרת מאסטר כנף -גנים אלה בונים כנפיים במקטעי בית החזה ובמבנים שונים, כמו מלכודות ג'ין, בקטעים אחרים. סוג הכנף - elytra, haltere, כנף מעופפת וכו 'תלוי גם במיקום הקטע בגוף ובמין בו הוא מתבטא. לדוגמה, לשפיריות יש שתי כנפיים מעופפות, ואילו הכנף הראשונה בחיפושית היא אליטרון מגן, והאגף השני בזבוב הוא איבר פרופריוספטיבי.
תכנות מסוג זה מכונה הומולוגיה סדרתית. נחזור למושג זה בהמשך, אך זהו רעיון חשוב מבחינה רעיונית ב- Evo-Devo. הומולוגים סדרתיים הם סוג מיוחד של הומולוג שבו אותו סוג של רקמה המבטאת קבוצת ליבה של מווסתים גנטיים (מארגן) נמצא במספר נקודות לאורך ציר הגוף. דוגמה אחת קלה להבנה להומולוגים סדרתיים הם ידיים ורגליים. מבחינה התפתחותית, הם כמעט זהים בחלק הראשון של צמיחתם, הם רק מגדילים הבדלים מהותיים מאוחר יותר. הם מבטאים את אותם גנים של איבר הליבה (כפי שנשקול בקרוב) ויש להם אותן עצמות, אם כי בצורות מעט שונות. הומולוגים סדרתיים חשובים להפליא בהבנתנו את האבולוציה מכיוון שהם "זולים" מבחינה גנטית, מכיוון שהם משתמשים באותה רשת רגולטורית של גנים, אך הם יכולים לספק פונקציות חדשות - כמו שימוש בכלים בתוספת תנועה. גורלו של הומולוג סדרתי תלוי לעתים קרובות בגנים של הוקס המתבטאים באזור המארגן ההומולוגי הסדרתי.
יציאות המארגן לא מושפעות רק מקוד ה- Hox, הן יכולות גם להגדיר בעצמן את קוד ה- Hox (כמו במקרה של Nanos ו- Bicoid) או יכולות להקים ציר אחר (כמו הציר הדורסוונטרלי של צפרדע או הציר האנטרופוסטריורי של גפה). כעת, לאחר שיש לך אחיזה איתנה ברשתות רגולטוריות של גנים, נוכל לבחון מקרוב את המארגן המפורסם שכבר שקלנו - מארגן ספמן-מנגולד של גסטרולה הצפרדע.
הגנטיקה של מארגן ספמן-מנגולד והנוטוכורד
אם אתה זקוק לרענון מהיר על מארגן ספמן-מנגולד, בדוק מחשוף וגסטרולציה. בקצרה, מארגן ספמן-מנגולד הוא מזודרם שנמצא במיקום הסהר האפור, הקוטב הגבי של עובר הצפרדע (או הניוט). מארגן זה, כשהוא מתפתח לנוטוכורד, גורם להיווצרות מבנים גביים כמו מערכת העצבים המרכזית ועמוד השדרה. דו-חיים עם מארגן ספמן-מנגולד נוסף מגדלים ציר A/P שני (ראשנים תאומים צמודים) ודו-חיים החסרים מארגן ספמן-מנגולד מתפתחים ל"חתיכת בטן".
בעזרת גנטיקה נוכל להסביר שתי שאלות ענק: 1) מה גורם למארגן להתפתח? ו 2) כיצד הוא גורם להיווצרות מבנים גביים? השלב הראשון ביצירת המארגן הוא סיבוב קליפת המוח עצמו, כפי שמוצג באיור 1 של סיבוב קליפת המוח. בסיבוב קליפת המוח, מיקרוטובולים מסובבים את קליפת המוח החיצונית של הביצית המופרית ביחס למסה הפנימית. לקוטב הצמחי יש כמה חלבונים מקומיים ו-mRNA הקשורים לשלד הציטו שלו. במסה הפנימית זה כולל VegT ו- Vg1 7,8. בקליפת המוח החיצונית יש חלבון Disheveled (Dsh) מקומי באופן צמחי, מרכיב של מסלול Wnt, המועבר לכיוון קוטב החיות במהלך סיבוב קליפת המוח 9. העקירה של Dsh יוצרת אזור חדש - חלק מהעובר עם Dsh, אך ללא Vg1 או VegT (איור 8) 10. Dsh עוזר למקם ולייצב b-catenin, גורם השעתוק של מסלול Wnt הקנוני. כעת יש לנו Vg1 ו- VegT בקוטב הצמחי ו- Dsh ו- b-catenin בקצה הגבי העתידי.
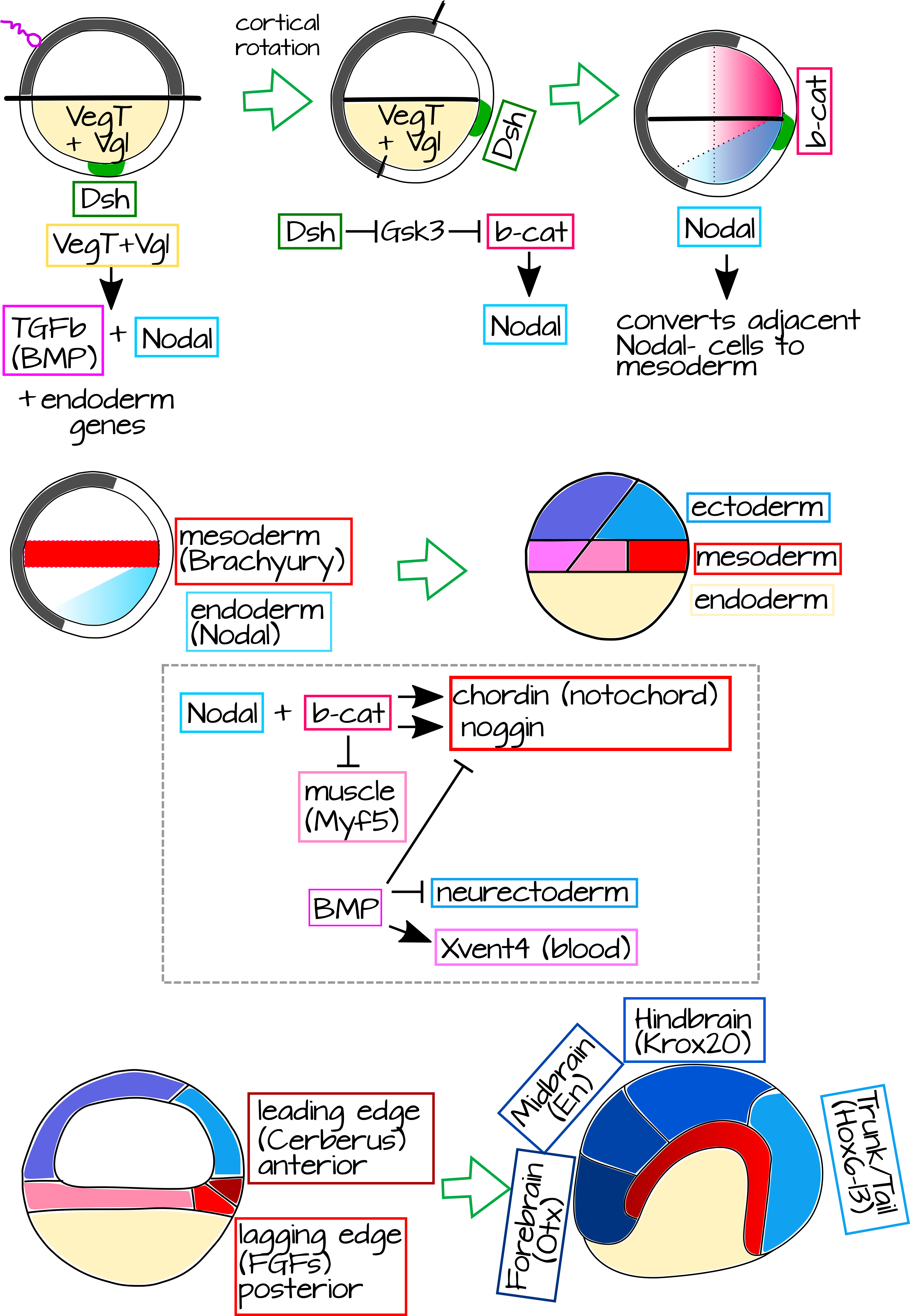 |
איור 8: גנטיקה של מארגן ספמן-מנגולד. חלבונים שמקורם באימהות מקימים ציר של בעלי חיים/צמחי בביצית הלא מופרית, כולל VegT ו- Vg1 בקליפת המוח הפנימית של הקוטב הצמחי (כתום). לאחר ההפריה, סיבוב קליפת המוח מביא את חלבון קליפת המוח החיצונית של הקוטב הצמחי Disheveled (Dsh, ירוק) למגע עם קליפת המוח הפנימית של בעלי החיים, מה שמניע את הצטברות ה-b-catenin בתאי הגב. חלבוני קוטב צמחי מפעילים את הביטוי של TgFb (BMP) ו-Nodal. Nodal ממיר את תאי ה-Nodal הסמוכים למזודרם (אדום), וכתוצאה מכך פס של מזודרם ממש מעל הקוטב הצמחי. בינתיים, b-catenin משפר את ביטוי הצמתים בחלק האחורי של העובר - וכתוצאה מכך ביטוי Nodal גבוה בקליפת המוח הפנימית הצמחית הגבית. זה, בתוספת b-catenin, מניע את הביטוי של גנים מארגנים כמו Chordin ו- Noggin, תוך דיכוי גורלות מזודרם גחון כמו שריר. Chordin ו-Noggin מציינים את ה-notochord, אשר יתארך ויגרום לאקטודרם הגבי (כל האקטודרם כחול) שנמצא מעליו כדי להפוך ל-neurectoderm. BMP מדכא את פעילות המארגן בצד הגחון של העובר, בפרט אקטודרם הגחון יהווה עור, לא neurectoderm. ל-notochord עצמו יש קצה מוביל (קדמי) ופיגור (אחורי) המבטא גנים שונים ויגרום לחלקים שונים של מערכת העצבים המרכזית. קובץ svg הניתן לעריכה של נתון זה זמין בכתובת https://scholarlycommons.pacific.edu/open-images/27/ |
ככל שהמחשוף מתקדם, VegT ו- Vg1 מעוררים את הביטוי של Nodal (במיוחד חלבון הקשור לצומת Xenopus או Xnr) בתאים הצמחיים ביותר. תאים אלה יהפכו לאנדודרם. Nodal, הומולוג של TgFb, מאותת לתאים סמוכים חסרי Nodal (תאי Nodal), תאים אלה יהפכו למזודרם. ביטוי נודלי נגרם גם על ידי b-catenin. זה יוצר שיפוע של Nodal שהוא גבוה יותר בצד הגבי, שם הוא מושרה על ידי VegT ו- Vg1 כמו גם b-catenin. בעוד b-catenin מיוצב בחלק רחב של רקמת הגב, ביטוי Nodal מוגבל לצד הצמחי של העובר שבו הוא מתבטא בכל רחבי, אבל הגבוה ביותר שבו b-catenin קיים (איור 8). כאשר איתות Nodal פוגש תאים מזודרמליים עם b-catenin גבוה, אנו רואים ביטוי של חבילת גנים חדשה - הגנים המארגנים Chordin, Noggin, Frizzled ועוד (איור 8) 11.
גנים מארגנים אלה הם גורמי שעתוק וחברים במסלולי העברת אותות. חלקם יציינו את המזודרם המארגן כ"גב "(notochord ו- somites) וחלקם יאותתו לאקטודרם המונח במהלך גסטרולציה ויציינו אותו כ"עצבי" ולא אפידרמיס. המכניקה הפיזית של תהליך זה מרתקת, ונוכל ללמוד אותם מאוחר יותר אם אתה מעוניין, אך לעת עתה אתמקד רק בגנטיקה ואסכם בקצרה את תנועות הרקמות. כפי שראית קודם, תאי מארגן מזודרמליים עוברים פיתול בשפה הגבית של הבלסטופור וזוחלים לאורך התאים האקטודרמליים המונחים כדי ליצור שכבה עבה מתחתיהם. רוב המזודרם בעובר הגסטרולציה מבטא BMP4, חלבון המופרש ממשפחת TgFb. BMP4 זה מאותת לאקטודרם שמעל להפוך לאפידרמיס. עם זאת, המארגן מבטא אנטגוניסטים BMP4, כמו Chordin ו Noggin 12. זה גורם לאקטודרם מעל המזודרם המארגן לקחת על עצמו גורלות עצביים.
המארגן עצמו אינו מבנה הומוגני, הוא מורכב מכמה אוכלוסיות תאים המבטאות גנים שונים של איתות וגורמי שעתוק. לדוגמה, הקצה המוביל של המארגן (החלק שייכנס ראשון ויהפוך לנוטוכורד הקדמי ביותר) מבטא אנטגוניסט Wnt ו- BMP4 בשם Cerberus 13. Cerberus וגנים מובילים אחרים מציינים את החלקים הקדמיים ביותר של הנוטוכורד באמצעות רשתות רגולטוריות של גנים שיובילו בסופו של דבר לביטוי של הגן הקדמי Otx2 14 באקטודרם שמעל. תאים ארגוניים העוקבים אחר קצה מוביל זה מבטאים את מולקולת האיתות המופרשת FGF, המניעה את הביטוי של גן המוח התיכון Krox20 באקטודרם שמעליו. גם Otx2 וגם Krox20 עוזרים ליצור את דפוס ביטוי הגנים של Hox, יחד עם גנים מארגנים אחרים המתבטאים בתבנית אנטרופוסטורית בנוטוכורד 14. בדרך זו, ה-notochord מסוגל לכוון את הדפוס האנטרופוסטורי של הצינור העצבי (איור 8).
מארגנים אחרים
ישנם המון מארגנים אחרים בביולוגיה התפתחותית ותכונותיהם הבסיסיות נותנות לנו תובנות לגבי הגנטיקה של האבולוציה. הם מוגדרים על ידי גורמים חיצוניים, למשל על ידי צומת של תאים המבטאים צמודי צומת ו- b-catenin במארגן Spemann-Mangold. הם פועלים על רקמות אחרות באמצעות מולקולות איתות מופרשות, למשל chordin מעכב BMP4 כדי לציין neurectoderm. ופעילותם על רקמות אחרות יכולה להשתנות לאורך זמן ו/או מרחב, למשל היכולת של הנוטוכורד לגרום למבנים של מערכת העצבים הקדמית והאחורית כאחד.
ל-notochord יש פעילות מארגנת שלישית על האקטודרם (שלא לדבר על פעילות מארגנת על המזודרם), זה דפוס של ציר D/V של הצינור העצבי. במהלך גסטרולציה, האקטודרם השוכב מעל הנוטוכורד מגיב לאותות ממנו להתקפל לצינור ושוקע מתחת לאפידרמיס. צינור זה מקוטב לצד הגבי (קרוב לאפידרמיס שמעל) ולצד הגחון (קרוב לנוטוכורד). אות Shh מהנוטוכורד מניע הפרשת Shh מהצד הגחון. שיפוע Shh זה מתנגד לשיפוע BMP (משפחת TGFb) הגבי שיזם איתות אפידרמיס (איור 9).
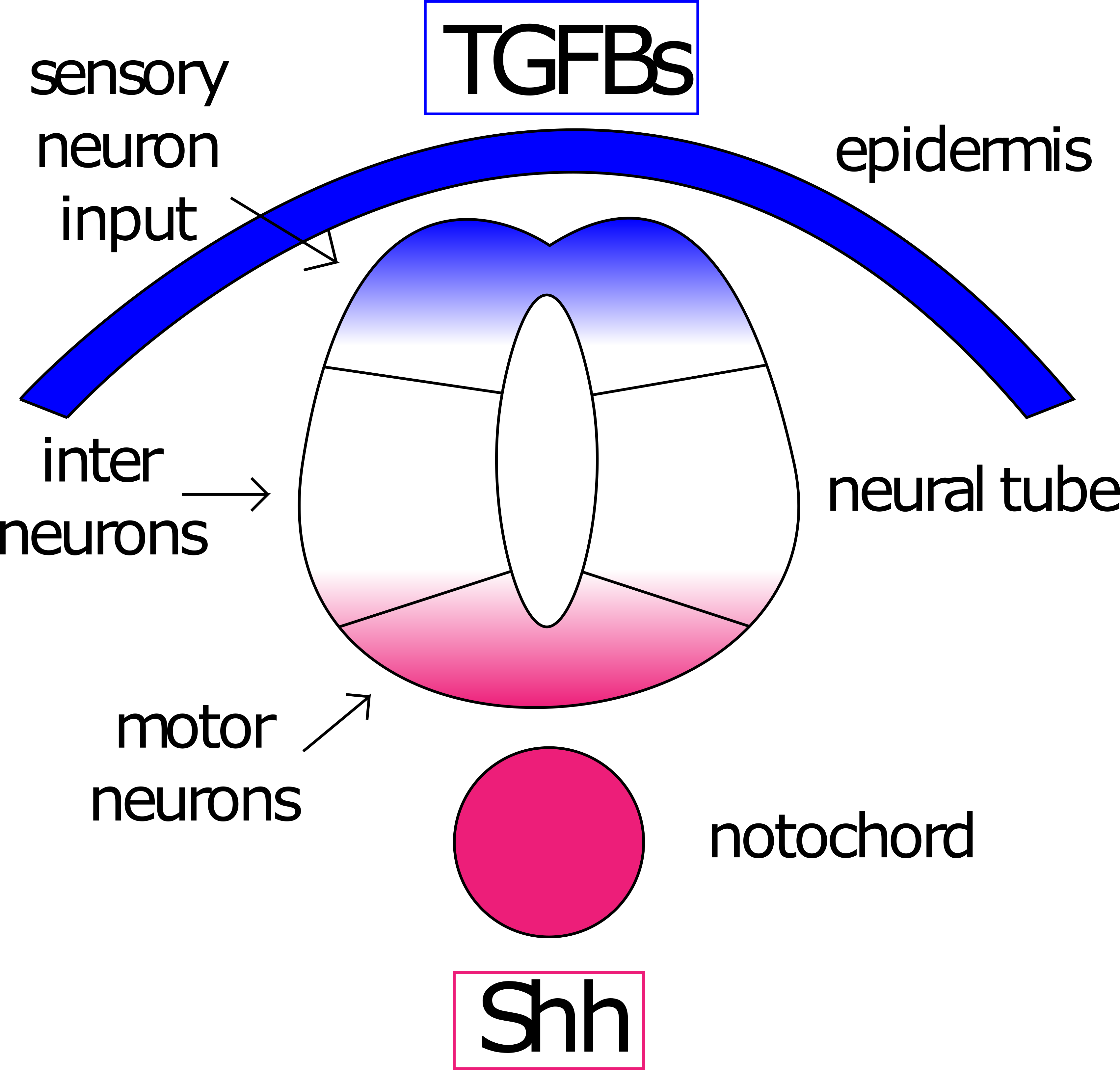 איור 8: שיפועים מנוגדים מדפיסים את הצינור העצבי. TGFBs (כחול) מהאפידרמיס וקיפוד סוניק (ששש, אדום) מתבנית הנוטוכורד הציר הגב/הגחון של הצינור העצבי. נוירונים שנחשפו לריכוז גבוה של TGFBs יהפכו לנוירונים תחושתיים, אלה שנחשפים לרמות גבוהות של Shh יהפכו לנוירונים מוטוריים. נוירונים שנחשפים לרמות נמוכות של שניהם יהפכו לאינטרנוירונים. דמות מאת אמנדה לו ואג'נה ריברה
איור 8: שיפועים מנוגדים מדפיסים את הצינור העצבי. TGFBs (כחול) מהאפידרמיס וקיפוד סוניק (ששש, אדום) מתבנית הנוטוכורד הציר הגב/הגחון של הצינור העצבי. נוירונים שנחשפו לריכוז גבוה של TGFBs יהפכו לנוירונים תחושתיים, אלה שנחשפים לרמות גבוהות של Shh יהפכו לנוירונים מוטוריים. נוירונים שנחשפים לרמות נמוכות של שניהם יהפכו לאינטרנוירונים. דמות מאת אמנדה לו ואג'נה ריברה
קובץ svg הניתן לעריכה של נתון זה זמין בכתובת https://scholarlycommons.pacific.edu/open-images/28/
שני שיפועים מנוגדים אלה מובילים לשיפועים של גורמי שעתוק פעילים המגיבים למסלולי העברת אותות אלה ולביטוי של גנים עם cis-אלמנטים רגולטוריים המגיבים לגורמי שעתוק אלה (איור 9) 15. Le Dreu ו- Marti מציינים 11 תחומים נפרדים המזוהים על ידי ביטוי ייחודי של גורם שעתוק ותת-סוג עצבי 15.
מארגן מפורסם נוסף הוא כזה שנתקלת בו Intro Bio: מארגן הגפיים הטטרפודים. זהו מארגן בן שני חלקים, עם קטע דק וארוך של תאים בנקודה המרוחקת ביותר של הגפה המבטאת ומפרישה FGFs מעודדי צמיחה, והנקודה האחורית ביותר של האיבר המבטאת את סוניק קיפוד (Shh, איור 10). חלק ה- FGF של הגפה נקרא רכס אקטודרמלי אפיקלי. מרכז האיתות הוא זה ששומר על הגפה להוסיף אלמנטים דיסטליים נוספים לקצה. באופן זה, היסודות הפרוקסימליים יותר, כמו עצם הזרוע והמרפק, מתפתחים תחילה. אלמנטים דיסטליים יותר, כמו האמה, פרק כף היד והידיים, מונחים מאוחר יותר. מעניין, נראה כי מערכת זו נמצאת על טיימר, במקום להיות בדוגמת מרחבית לחלוטין. אם חלוקת התא מעוכבת באמצע התפתחות הזרוע, רדיוס, אולנה והומרוס יכולים להיות קטנים או נעדרים, עם חגורת יד וכתף פרופורציונלית בדרך כלל. זה נראה קצת דומה למוטנט פער ב-Drosophila, אם כי מנגנון ההתפתחות שונה לחלוטין.
איור 10: מארגן הגפיים הטטרפודים. הרכס האקטודרמלי האפיקי (AER, אדום) מפריש FGFs השומרים על התפשטות התאים הבסיסיים ושומרים על איתות Shh מאזור הפעילות המקוטבת (ZPA, כחול). חלבון Shh המופרש מתאי ZPA יוצר שיפוע לאורך ציר A/P (אגודל\ ורוד) של הגפה. נמוך/לא Shh מציין גורלות של האגודל והאצבע המורה, Shh גבוה מציין גורלות של ורוד ואצבע. מה קורה אם נוסיף ששש לצד השני של הגפה המתפתחת? תמונה ממשתמש ויקימדיה פיטרוץ תחת רישיון CC BY-SA 3.0
מרכז האיתות Shh הוא מארגן אמיתי. זה לא רק מפריש מולקולת איתות כמו ה- AER, אלא שמולקולת האיתות הזו פועלת בצורה מדורגת. רמות גבוהות של Shh גורמות להתפתחות של אלמנטים אחוריים של הגפה, כמו הרדיוס או הזרת, בעוד שרמות נמוכות גורמות להתפתחות של אלמנטים אמצעיים, כמו האצבעות האמצעיות, והיעדר Shh מוביל לאלמנטים קדמיים, כמו אגודל.
באופן כללי, התמרת אותות ומפלי גורמי שעתוק אחראים לחלק גדול מהתבניות של העובר המתפתח. זה נותן לנו רמז גדול כיצד האבולוציה יכולה לפעול על התפתחות לשינוי מורפולוגיה של מבוגרים. לשינויים קטנים בכמות או במיקום של מורפוגן יכולים להיות השפעות גדולות על המבוגר. אותה "ערכת כלים" של גנים נשמרת באבולוציה, אך נעשה בה שימוש חוזר גם בהקשרים רבים ושונים. למשל תפקידו של Shh בדפוס מערכת העצבים המרכזית שלך כמו גם בידיים שלך. שימוש חוזר זה באלמנטים של ארגז כלים בהקשרים חדשים מעניק להם פונקציות חדשות - בהתאם למפעיל המקומי ולמדכאים שנמצאים שם, הגנים של מסלול העברת האותות הזמינים, כמו גם שינויים כרומוזומליים אפיגנטיים שכבר קיימים. עד כה התמקדנו בעיקר בשיפועים ובמארגנים בדפוסים, אך בדפוסים נבחן כמה דרכים אחרות לדפוס עובר.