
3.1: המקרה המוזר של Bicoid: מודלים להכנת שיפוע - מודל עולה
- Page ID
- 207543

מודלים להכנת שיפוע
שיפועים מורפוגניים חשובים להפליא בפיתוח עבור רקמות דפוסיות. אם רקמה הומוגנית לחלוטין (כל תא מבצע את אותה משימה) היא לא יכולה להיות מורכבת ולעתים קרובות אינה עמידה בפני תנודות סביבתיות. במקרה של Bicoid in Drosophila, אנו שוקלים שיפוע מורפוגני המדפיס את הציר האנטרופוסטורי - כאמור במבוא, יותר מדי Bicoid נותן לנו ראש ענק ומעט מדי לא נותן לנו ראש בכלל. ישנם איפשהו בסביבות 60 גנים שונים המגיבים לרמות שונות של חלבון ביקואידי ואחד המוקדמים שהתגלו הוא הגיבן. הגיבן נותן מידע מיקום לעובר זבוב, הוא מחדד את הגבול בין הקדמי לאחורי הזבוב. בעוד שיפוע החלבון הביקואידי רחב מאוד ודומה לשיפוע דיפוזיה, שיפוע הגיבן חד, עם רמות חלבון שונות באופן משמעותי בתוך 10 מיקרומטר, או כ-2 גרעינים 1.
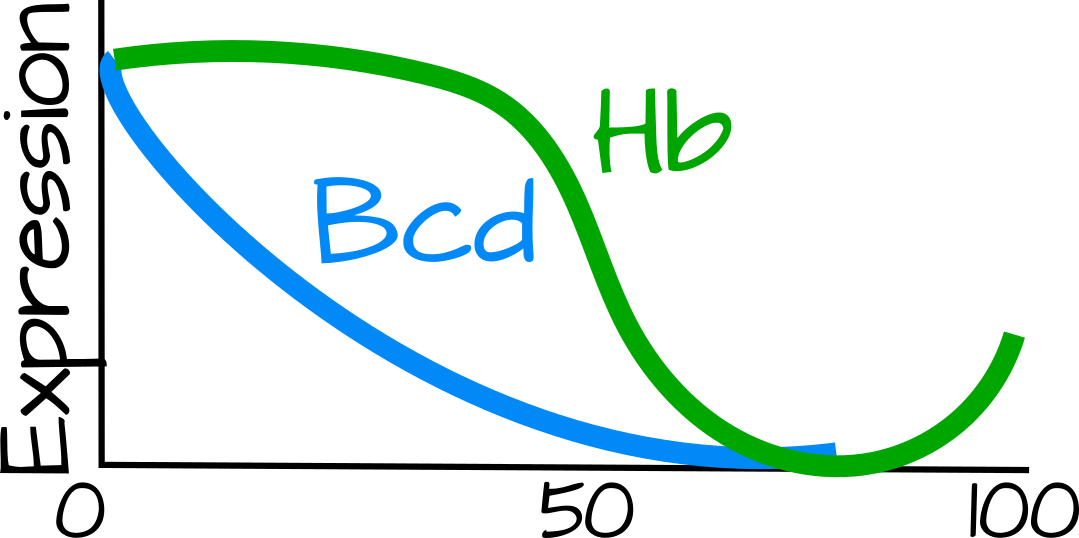
ברור שרמות הביקואיד חייבות להיות חשובות מאוד מכיוון ששיפוע הביקואיד והקריאה שלו (למשל רמות הגיבן) מדויקים להפליא. מודל מוקדם לשיפוע הביקואידי שנצפה ב-Drosophila הוא מודל ה-SSD (סינתזה, מומס, דיפוזיה), שבו מקור נקודתי לחלבון ביקואידי (מתורגם מ-mRNA דו-קואידי קדמי) מגדיר את שיפוע הריכוז. מודל ה- SDD הקשור (סינתזה, דיפוזיה, פירוק) משלב פירוק לאורך זמן של חלבון Bicoid, ומונע מהשיפוע להגיע לקצה האחורי של העובר. הוצעו מספר מודלים אחרים שיאפשרו תנודות בגודל העובר אך עדיין יובילו לקריאה נכונה של Bicoid:
- יותר mRNA/חלבון בעוברים גדולים יותר: זה יאריך את השיפוע עוד יותר בעוברים גדולים יותר.
- הסעות גרעיניות: זהה למודל SDD, אך חלבון Bicoid נקלט על ידי גרעינים כשהוא מתפזר על ידם. לפי מודל זה, ככל שהגרעינים ארוזים בצפיפות רבה יותר (עובר קטן יותר) כך החלבון הביקואידי נשלף מהר יותר מהציטופלזמה הסינציאלית. בעוברים גדולים יותר, החלבון יכול להתפזר רחוק יותר.
- מודל ARTS (mRNA פעיל, תחבורה, סינתזה): שלד הציטוס מעביר באופן פעיל mRNA Bicoid ליצירת שיפוע mRNA שמקבל קריאה כשיפוע חלבון כפי שהוא מתורגם. בעוברים גדולים יותר, שלד הציטוס יכול להעביר את ה-mRNA הלאה.
- דיפוזיה ופירוק mRNA: זה דומה למודל ARTS, אלא שבמודל זה mRNA Bicoid מתפרק לאורך זמן כדי לשמור על שיפוע mRNA. לזבובים גדולים יותר יכולים להיות שיעורי פירוק נמוכים יותר של mRNA Bicoid או הובלה מהירה יותר.
- הקלה על דיפוזיה של חלבון. זה דומה למודל ARTS, רק שהוא כרוך בהובלה של חלבון Bicoid על ידי שלד הציטוס. במקרה זה, זבובים גדולים יותר יכולים לקבל הובלה מהירה יותר של חלבון Bicoid.
ישנן עדויות בעד ונגד כל אחד מהמודלים הללו, דבר המצביע על כך ששיפוע החלבון הביקואידי נשמר ככל הנראה על ידי שילוב של מודלים אלה. בנוסף, קריאת חלבון Bicoid עשויה להיות מורכבת יותר ממה שאנו חושבים - חלבונים מרובים עשויים להשפיע על האופן שבו Bicoid מתקשר עם המטרות שלו.
עדויות בעד ונגד הדוגמניות
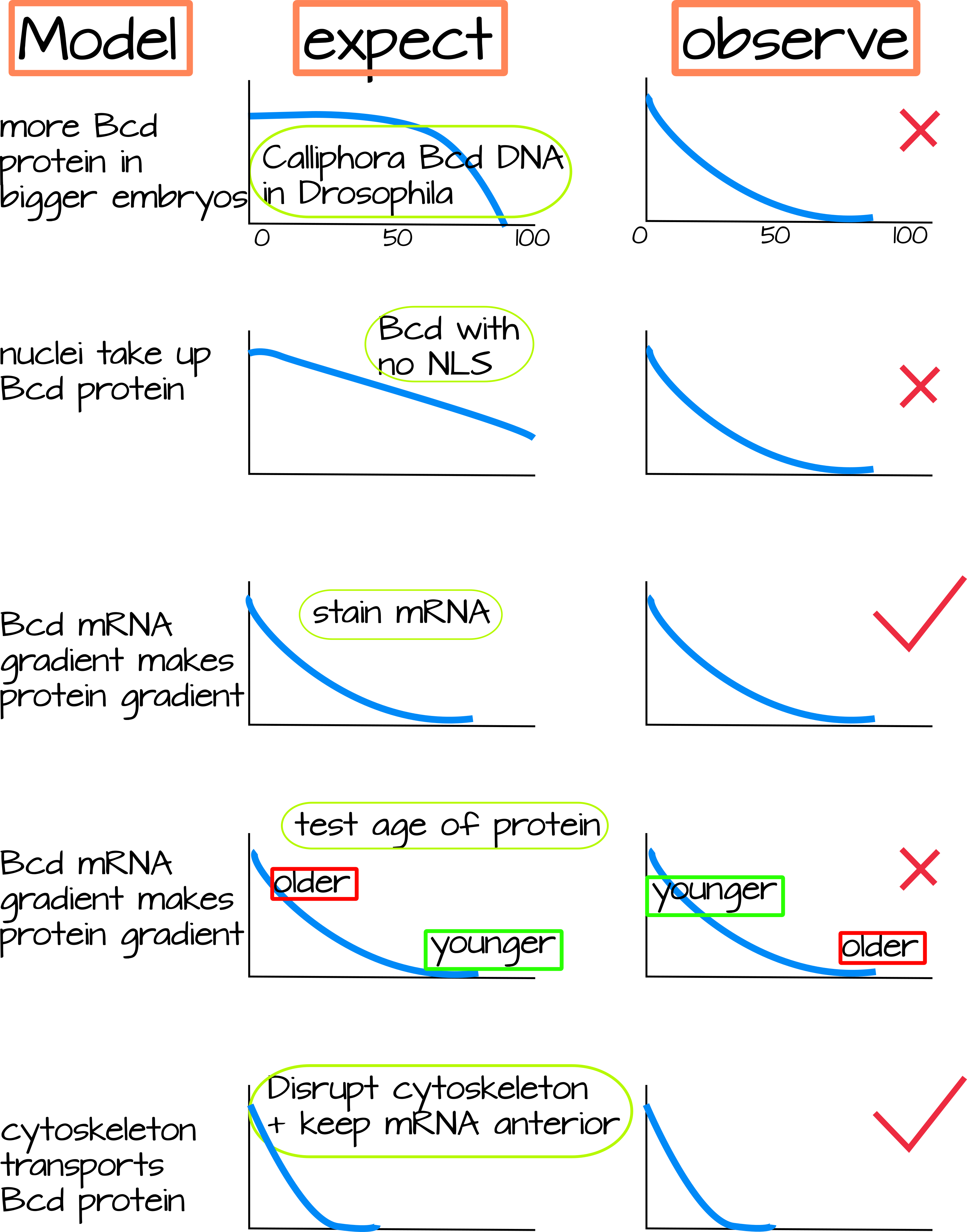
נתונים הן של תסיסנית והן מדיפטרנים אחרים, גדולים יותר, סיפקו ראיות התומכות בכל אחד מהמודלים המפורטים לעיל. בדיקות נוספות זייפו את התחזיות של חלק מהמודלים, וזה הנושא של סעיף זה. להלן אני מתאר כל דגם ומבחן של אותו מודל, אני כולל גם דמות הממחישה למה המודל מצפה ומה נצפה בפועל. אני ממליץ על סוג זה של דמות סופר-רדוקציוניסטית כדי לעזור לסכם מערך נתונים גדול, אך עלינו תמיד לזכור שהוא לא מספר את כל הסיפור.
1. מודל חלבון נוסף. עובר יכול לקבל יותר חלבון על ידי הגדלת כמות ה- mRNA, הגדלת קצב התרגום של אותו mRNA, או הפחתת קצב פירוק החלבון. תומך בכך הוא הממצא כי הכנסת עותקים נוספים של Bicoid (יותר mRNA ויותר חלבון) לזבוב פירות מגדילה את שיפוע הביקואיד ואת גודל הראש. כדי לראות אם זה מה שזבובים עם עוברים גדולים יותר עושים כדי ליצור שיפוע גדול יותר, החוקרים שמו DNA Bicoid, עם cis-אתרים רגולטוריים ו-UTRs, מזבובי עוברים גדולים (Calliphora) ב-Drosophila. הם גילו שהגן הביקואידי מעוברים גדולים לא עשה שיפוע ארוך יותר ב תסיסנית. לכן, אם ישנם גורמים המשפיעים על יציבות ה-mRNA או פירוק החלבון, הם חייבים להיות ספציפיים לזבובים הגדולים 3.
2. הסעות גרעיניות. ניסויים מוקדמים בנושא זה העלו כי הוספת אות לוקליזציה גרעינית לחלבון Bicoid עשתה שיפוע רדוד יותר 3. ניסויים מאוחרים יותר בדקו זאת על ידי הכנת תסיסנית עם גרסה מוטנטית של Bicoid שלא הצטברה בגרעינים. אם יבוא גרעיני הפך את שיפוע החלבון הביקואידי לתלול יותר, אז הם היו רואים שיפוע רחב יותר לאחר עיכוב יבוא גרעיני. עם זאת, הם ראו שיפוע חלבון ביקואידי למראה רגיל למדי, וטענו בתוקף נגד מודל זה 4.
3 ו-4. מודלים של שיפוע mRNA. בשני הדגמים 3 ו-4, mRNA מועבר באופן פעיל סביב התא על ידי שלד הציטוס. זה מנוגד ישירות למודל SDD, שבו חלבון Bicoid מתורגם מ-mRNA Bicoid קשור קדמית. בשני מודלים אלה של שיפוע mRNA, שיפוע החלבון הוא פשוט "קריאה" של שיפוע ה- mRNA. עדויות לשני המודלים הללו מגיעות ממדידה מדוקדקת של mRNA Bicoid בפיתוח עוברים (איור 2). מדידות זהירות אלה מראות ש-mRNA Bicoid עצמו יוצר שיפוע, ואינו רק מקור נקודתי 5. תנועתו תלויה בשלד הציטוס, אם שלד הציטוס מופרע, שיפוע ה-mRNA מופרע גם הוא 6. עדויות נגד מודל זה נובעות מהממצא שחלבון Bicoid בחלק האחורי של העובר מבוגר יותר מחלבון Bicoid בחלק הקדמי. אם mRNA Bicoid מתפזר לכיוון האחורי כדי ליצור שיפוע, אז חלבון Bicoid עשוי מאותו mRNA, היינו מצפים לראות חלבון Bicoid צעיר יותר בחלבון Bicoid בגיל האחורי והמעורב בקצה הקדמי. עם זאת, Durrieu et al מצאו את ההפך, מה שמרמז שלמרות ש-mRNA Bicoid עשוי ליצור שיפוע, זה לא הקובע העיקרי של שיפוע החלבון 7.
5. דיפוזיה חלבונית מקלה. שיבוש שלד הציטוס על ידי טיפול בעובר תסיסנית בתרופות אנטי-אקטין או אנטי-מיקרוטובוליות מוביל לשיבוש שיפוע החלבון. מכיוון שקשירת mRNA בקוטב הקדמי תלויה גם במיקרוטובולים, רק שיבוש שלד הציטוס לא יגיד לך אם הובלת mRNA או הובלת חלבון היא שמובילה לשיפוע החלבון. מחקר אחד השתמש בטריק מצחיק - אם תגדל עוברי תסיסנית בתנאים דלי חמצן (היפוקסי), ה-mRNA הביקואידי יישאר בקצה הקדמי גם אם המיקרו-צינורות יופרעו. מחקר זה מצא שחלבון Bicoid עדיין יוצר שיפוע גם אם mRNA Bicoid לא עושה זאת, וכי שיפוע זה תלוי בשלד הציטו 8.
מודל עולה
למרות כל הנתונים הסותרים לכאורה, מתחיל לצוץ מודל שמסביר את החוסן של שיפוע החלבון הביקואידי ומסביר כיצד השיפוע עצמו יכול להתרחב לגופים גדולים או קטנים יותר. ראשית, mRNA Bicoid מנותק ברובו בקצה הקדמי של העובר, אך גם עובר לאורך קליפת המוח (הקצה החיצוני) של העובר באמצעות מיקרוטובולים. חלבון ביקואידי מתורגם מה- mRNA Bicoid, כאשר כמות תרגום גבוהה יותר מתרחשת בקצה הקדמי, שם ריכוז ה- mRNA הביקואידי גבוה יותר. סביר להניח שחלק מהחלבון הביקואידי מתורגם גם מהריכוז הנמוך יותר של mRNA Bicoid שנע מאחור דרך מיקרוטובולים. זה מוביל לשיפוע של חלבון Bicoid שהוא מעט רחב יותר ממה שהיינו מצפים לו היה מקור נקודתי פשוט לחלבון Bicoid (כלומר תרגום מקוטב קדמי Bicoid mRNA).
שיפוע חלבון זה משתנה גם כאשר חלבון Bicoid מועבר סביב העובר על ידי השלד (באמצעות חוטי אקטין ומיקרוטובולים). בדרך זו, יש שליטה על השיפוע על ידי שלד הציטוס ועל ידי החלבונים המתווכים אינטראקציה בין Bicoid (חלבון ו- mRNA) לבין שלד הציטוס. חלבונים מתווכים אלה עשויים להיות רגישים לרמות הביקואידיות המקומיות ועשויים לאפשר לביקואיד לנוע מהר יותר או לאט יותר בהתאם למיקום האנטרופוסטורי ולרמת השיפוע. לדוגמה, זבוב גדול עשוי להכיל פחות חלבון ביקואידי על ידי דיפוזיה ב -15% מאורכו (EL) מאשר לזבוב קטן ב -15% מאורכו. עם זאת, כל החלבון הביקואידי ב -15% EL בזבוב הגדול יכול להיספג על ידי החלבונים המתווכים ולעבור אחורה כדי להרחיב את השיפוע. מצד שני בזבוב קטן, הכמות הגדולה יותר של חלבון Bicoid ב -15% EL עלולה להרוות יתר על המידה את החלבון המתווך ולהעביר אותה בצורה פחות יעילה.
בנוסף, חלבונים ביקואידים ממינים שונים של זבובים ידועים כבעלי תכונות מעט שונות, למשל Bicoid ממיני זבובים גדולים יותר כגון Calliphora vicinia ו Lucilia sericata אינם מסוגלים להציל לחלוטין מוטציות Drosophila Bicoid 3. תכונות שונות אלה יכולות גם למלא תפקיד באינטראקציות חלבון-חלבון המערבות את שלד הציטוס. לבסוף, החלבונים המתווכים בין המינים יכולים גם הם להיות שונים, חלקם עשויים להיקשר לביקואיד ביעילות רבה יותר ו/או לנוע לאורך השלד בצורה יעילה יותר.