
1.3: בקצרה, גנטיקה
- Page ID
- 207461

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)כמה מונחים בסיסיים
DNA הוא פולימר חומצת גרעין כפול גדילי שהוא החומר התורשתי העיקרי ברוב האורגניזמים החיים. כל קווצת DNA מורכבת מנוקלאוטידים הקשורים קוולנטית. כל נוקלאוטיד מורכב מסוכר, פוספט ובסיס חנקני. הקשר הקוולנטי בין נוקלאוטידים מתרחש בין הסוכר של נוקלאוטיד אחד לפוספט של הבא. ב- DNA יש סוג יחיד של סוכר אך 4 סוגים של בסיסים. בסיסים אלה - אדנין, תימין, ציטוסין וגואנין - נותנים לכל נוקלאוטיד את שמו (מקוצר A, T, C ו- G).
שני גדילי ה-DNA קשורים זה לזה על ידי קשרי מימן בין בסיסים חנקניים. לכל בסיס חנקני יש בסיס "משלים". זוגות עם T ו- C זוגות עם G אם אנו חושבים על גדיל DNA בעל כיוון של סוכר-פוספט-סוכר-פוספט-סוכר-סוכר-פוספט אז לגדיל השני של המולקולה יש כיוון של פוספט-סוכר-פוספט-סוכר-פוספט-סוכר. אנו מכנים סוג זה של אוריינטציה אנטי מקבילה ואנו מתייחסים לשני הגדילים כתוספים הפוכים. השלמה הפוכה מתייחסת הן לכיוון ההפוך שלהם (הפוך) והן לתבנית מליטה A = T G = C (השלמה).
ל-RNA יש מבנה דומה, אם כי הוא נמצא בדרך כלל כגדיל בודד. הוא מסוגל ליצור גדילים כפולים (או אפילו מבנים מורכבים יותר, למשל tRNA) באמצעות קשר מימן עם רצפי השלמה הפוכים. זה יכול לקשר מימן עם עצמו או עם מולקולות RNA אחרות.
הדוגמה המרכזית של הביולוגיה המולקולרית
לרוב כאשר אנו חושבים על גנטיקה מולקולרית אנו חושבים לייצר חלבונים פונקציונליים מגן המקודד ב- DNA, אם כי חלק מה- RNA מתפקד בפני עצמו.
ייצור החלבון מקוד DNA מתרחש באמצעות שני תהליכים מווסתים מאוד: שעתוק משתמש ב-RNA פולימראז כדי ליצור מולקולת mRNA חד-גדילית מגדיל אחד של מולקולת DNA כפולה. תרגום משתמש בריבוזום כדי ליצור פפטיד (חלק מחלבון או כולו) מה-mRNA (איור 1).
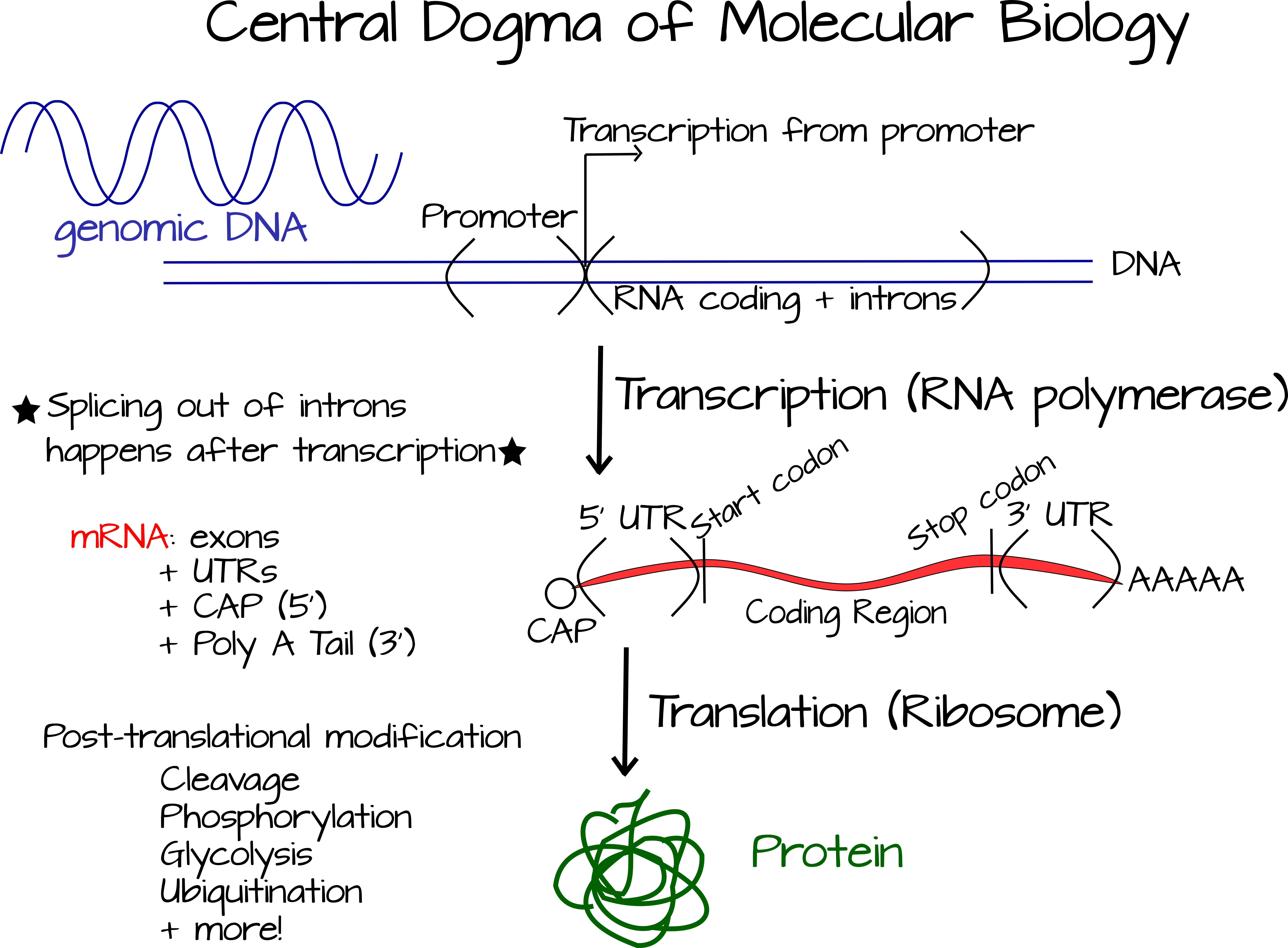
הערה צדדית מהירה מכיוון שאנשים רבים מבולבלים מכמה מהמונחים בהם אנו משתמשים בגנטיקה, אבולוציה והתפתחות. גן מתייחס לפיסת DNA המקודדת ל- RNA. RNA זה הוא לעתים קרובות mRNA אך יכול להיות גם tRNA, rRNA, snRNA, miRNA או סוג אחר של RNA "לא מקודד". mRNA הוא RNA המקודד לחלבון. רוב הזמן "ביטוי גנים" מתייחס לייצור mRNA ו/או החלבונים שהם מקודדים להם. ביטוי גנים הוא תהליך השימוש ב- DNA לייצור מוצר פונקציונלי. כפי שאתה בוודאי יודע, רוב התאים בגופך זהים גנטית (פחות או יותר); הם "חולקים גנום". עם זאת, לתאים שלך יש פונקציות שונות מאוד. פונקציות שונות אלה מונעות על ידי סביבות שונות ופרופילי ביטוי גנים שונים - כלומר התאים מבטאים גנים שונים. לדוגמה, אנו עשויים לצפות למצוא הרבה חלבון מיוסין בתאי שריר אך הרבה חלבון אקוופורין בתאי הכליה.
ויסות ביטוי הגנים (אפיגנטיקה)
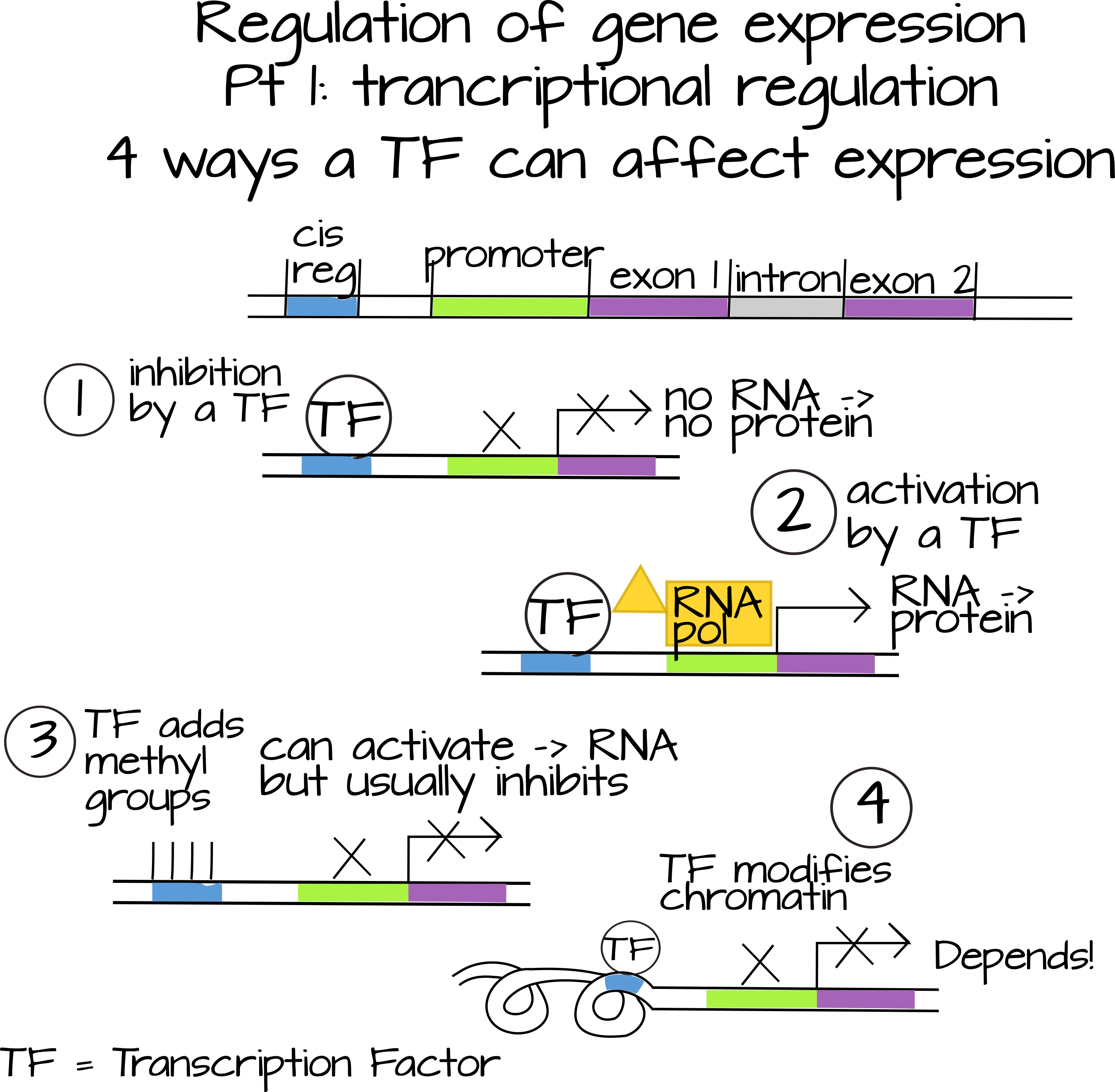
ביטוי הגנים מוסדר בשלוש רמות עיקריות (שניתן לחלק אותן להרבה תת-רמות רבות). ניתן לווסת את התעתיק עצמו (לעתים קרובות אנו מתייחסים לזה כאל הפעלת גן "או" כבוי "). ויסות התעתיק הוא תפקידם של גורמי שעתוק, הידועים גם כגורמים "פועלים טרנס". אלו הן מולקולות ניתנות לפיזור, לרוב חלבונים, הפועלות בדרכים רבות ושונות כדי להגביר או להוריד את קצב התעתיק של גן מסוים. גורמי שעתוק מסוימים נקשרים ישירות ל- cis-רצפים רגולטוריים, פיסות DNA בדרך כלל ליד אזור הקידוד של הגן, ומשפיעים על יכולתו של RNA פולימראז להיקשר למקדם הגן. הם יכולים לעשות זאת על ידי אינטראקציה ישירה עם חלבונים המעורבים בגיוס RNA פולימראז (למשל TFIID ו- Mediator), או על ידי שינוי הנגישות של האמרגן (למשל, חסימה פיזית של האמרגן, הוספה או הסרה של קבוצות מתיל, או אינטראקציה עם היסטונים ו חלבונים קושרים היסטון). גורמי שעתוק מסוימים נקשרים לגורמי שעתוק אחרים ומשפיעים על יכולתם לקשור DNA.
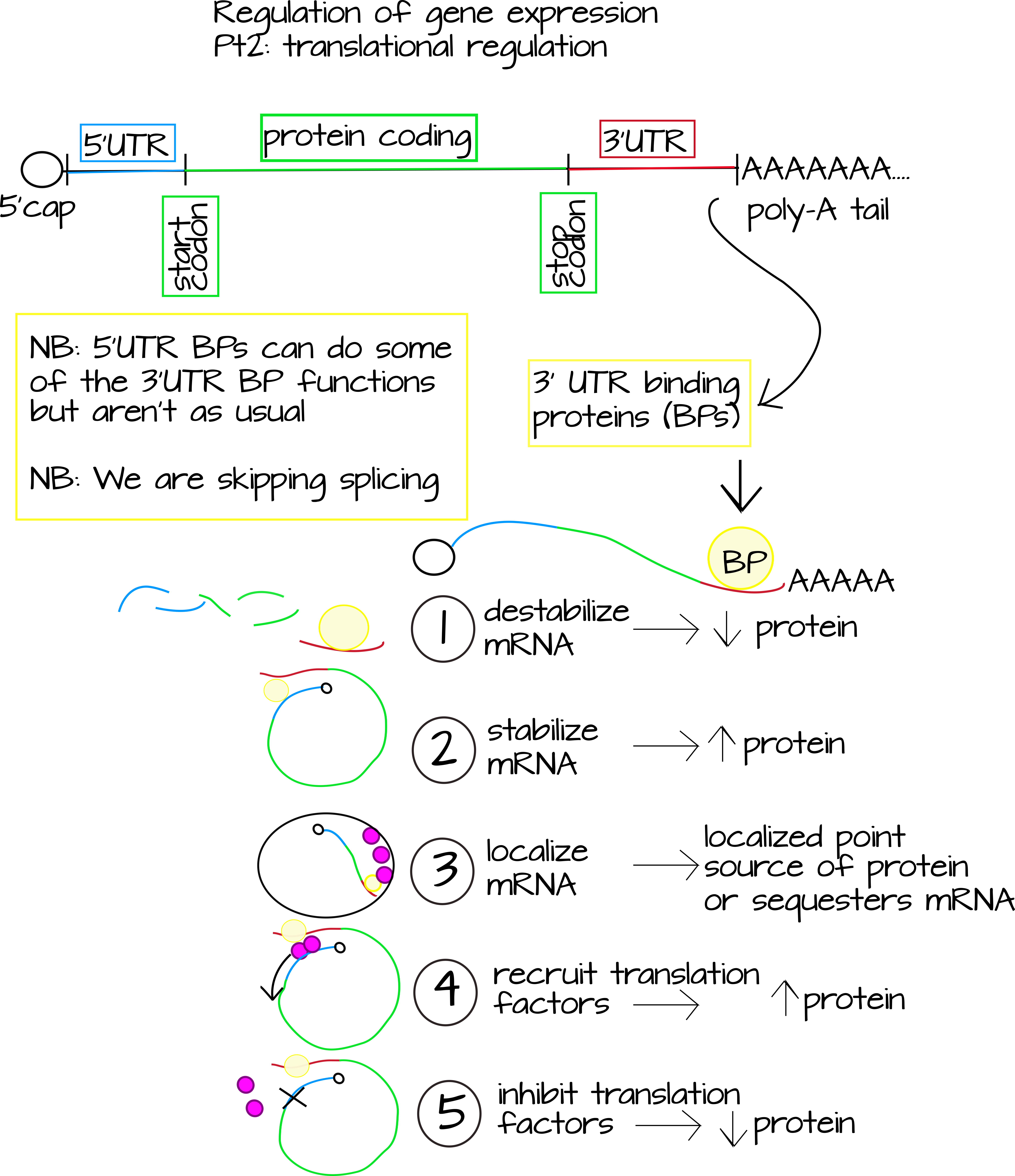
הרמה העיקרית השנייה של ויסות ביטוי גנים היא ויסות תרגום. זה כולל כל סוג של רגולטורים המשפיעים על כמות החלבון/פפטיד המיוצר על ידי מולקולת mRNA אחת. ישנן שתי דרכים שנחקרו היטב שזה קורה. הראשון הוא באמצעות הפרעות RNA (RNAi), שאולי שמעתם עליהן. RNAi משתמש בחתיכות קצרות של RNA לא מקודד שהן השלמות הפוכות מדויקות או קרובות מאוד לחלק מרצף mRNA. זה מכוון ל- mRNA להשמדה או חוסם תרגום. 1 הדרך השנייה הנחקרת היטב לווסת את התרגום היא באמצעות חלבונים קושרים ל-RNA. חלבונים אלה ייקשרו לעתים קרובות לרצפים ספציפיים הנמצאים מחוץ לאזור המקודד לחלבון ב- mRNA. אזורים אלה נקראים האזורים הלא מתורגמים או "UTRs". חלבונים שנקשרים כאן יכולים
- לערער את ה-mRNA, להוריד את רמות התרגום.
- לייצב את ה-mRNA, להעלות את רמות התרגום.
- מקם את ה- RNA לחלק מסוים של התא.
- חסום או מעכב את הקישור של גורמי תרגום, והוריד את רמות התרגום.
- לגייס גורמי תרגום, העלאת רמות התרגום. 2
הרמה העיקרית השלישית והאחרונה של ויסות ביטוי הגנים היא הרמה שלאחר התרגום. רמה זו מקיפה את רוב מה שאנו חושבים עליו כמסלולי העברת אותות, או אינטראקציות חלבון-חלבון וחלבון-מולקולות קטנות המשפיעות על יציבות ותפקוד החלבון. זה יכול לכלול גם שינוי קוולנטי של החלבון (לדוגמה, זרחון, דה-פוספורילציה, גליקוזילציה או מחשוף פרוטאוליטי) כמו גם שינוי לא קוולנטי (למשל עיכוב או הפעלה אלוסטרית, היווצרות הומו-והטרודימר). 3 אם אינך בטוח מה המשמעות של מונחים אלה כרגע, זה בסדר - את רובם לא נכסה ואת אלה שאנו מכסים קל יותר להבין בהקשר כאשר אתה לומד על מסלולי העברת אותות.
הפניות
- "מנגנוני השתקת גנים על ידי RNA דו-גדילי", 2004, טֶבַע, גונטר מייסטר ותומס טושל, דוי:10.1038/טבע02873
- "עולם חדש ואמיץ של חלבונים המחייבים RNA", 2018, ביקורות טבע: ביולוגיה מולקולרית של תאים, מתיאס וו הנצה, אלפרדו קסטלו, תומאס שוורצל ותומס פרייס דוי:10.1038/nrm.2017.130
- "שלושה הרגלים של מסלולי איתות יעילים ביותר: עקרונות של בקרת תעתיק על ידי איתות תאים התפתחותיים", 2002, גנים ופיתוח, סקוט ברולו וג'יימס וו פוסקוני, 10.1101/gad.976502