
11.7: Udhibiti wa Gene - Nadharia ya Operon
- Page ID
- 174703

Malengo ya kujifunza
- Kulinganisha watu inducible na watu repressible
- Eleza kwa nini udhibiti wa operons ni muhimu
Kila kiini cha nucleated katika kiumbe cha multicellular kina nakala za DNA sawa. Vilevile, seli zote katika tamaduni mbili safi za bakteria zilizoingizwa kutoka koloni moja kuanzia zina DNA hiyo, isipokuwa mabadiliko yanayotokana na mabadiliko ya hiari. Ikiwa kila kiini katika kiumbe cha multicellular kina DNA sawa, basi ni vipi seli katika sehemu tofauti za mwili wa viumbe zinaonyesha sifa tofauti? Vilevile, ni vipi kwamba seli moja za bakteria ndani ya tamaduni mbili safi zilizo wazi kwa hali tofauti za mazingira zinaweza kuonyesha fenotipu tofauti? Katika matukio hayo yote, kila kiini kinachofanana na jeni haina kugeuka, au kuelezea, seti sawa ya jeni. Subset tu ya protini katika seli kwa wakati fulani inaelezwa.
DNA ya jenomu ina jeni zote mbili za kimuundo, ambazo zinajumuisha bidhaa zinazotumika kama miundo ya seli au enzymes, na jeni za udhibiti, ambazo zinasimisha bidhaa zinazodhibiti usemi wa jeni. Maneno ya jeni ni mchakato uliowekwa sana. Ingawa kusimamia usemi wa jeni katika viumbe vya seli mbalimbali huruhusu upambanuzi wa seli, katika viumbe single-seli kama prokaryotes, kimsingi huhakikisha kwamba rasilimali za seli hazipotewi na kutengeneza protini ambazo seli hazihitaji wakati huo.
Elucidating utaratibu wa kudhibiti jeni kujieleza ni muhimu kwa uelewa wa afya ya binadamu. Vikwazo katika mchakato huu kwa wanadamu husababisha maendeleo ya kansa na magonjwa mengine. Kuelewa mwingiliano kati ya usemi wa jeni wa pathojeni na ule wa mwenyeji wake wa binadamu ni muhimu kwa kuelewa magonjwa fulani ya kuambukiza. Udhibiti wa jeni unahusisha mtandao tata wa mwingiliano ndani ya kiini kilichopewa kati ya ishara kutoka mazingira ya seli, kuashiria molekuli ndani ya seli, na DNA ya seli. Uingiliano huu unasababisha usemi wa jeni fulani na ukandamizaji wa wengine, kulingana na mazingira.
Prokaryotes na eukaryotes hushiriki baadhi ya kufanana katika utaratibu wao wa kudhibiti usemi wa jeni; hata hivyo, kujieleza kwa jeni katika eukaryotes ni ngumu zaidi kwa sababu ya kujitenga kwa muda na anga kati ya michakato ya transcription na tafsiri. Hivyo, ingawa udhibiti mkubwa wa kujieleza jeni hutokea kwa njia ya udhibiti wa transcriptional katika prokaryotes, udhibiti wa kujieleza jeni katika eukaryotes hutokea katika ngazi ya transcriptional na baada ya transcriptionally (baada ya nakala ya msingi imefanywa).
Prokaryotic Gene Kanuni
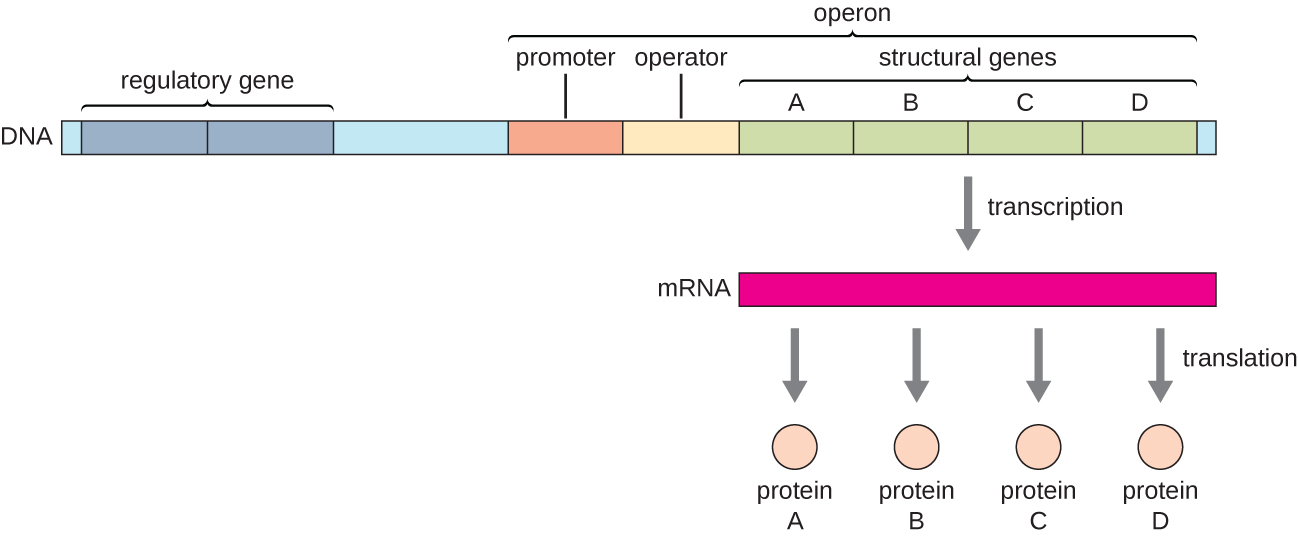
Katika bakteria na archaea, protini za kimuundo na kazi zinazohusiana ni kawaida encoded pamoja ndani ya genome katika block inayoitwa operon na ni transcribed pamoja chini ya udhibiti wa promota moja, na kusababisha malezi ya nakala polycistronic (Kielelezo\(\PageIndex{1}\)). Kwa njia hii, udhibiti wa transcription ya jeni zote za kimuundo encoding enzymes ambazo huchochea hatua nyingi katika njia moja ya biochemical inaweza kudhibitiwa wakati huo huo, kwa sababu watakuwa ama wote kuhitajika kwa wakati mmoja, au hakuna itahitajika. Kwa mfano, katika E. koli, jeni zote za kimuundo ambazo zinajenga enzymes zinahitajika kutumia lactose kama chanzo cha nishati ziko karibu na kila mmoja katika operon ya lactose (au lac) chini ya udhibiti wa promota moja, mtangazaji wa lac. Wanasayansi wa Kifaransa François Jacob (1920—2013) na Jacques Monod katika Taasisi ya Pasteur walikuwa wa kwanza kuonyesha shirika la jeni za bakteria kuwa operoni, kupitia masomo yao juu ya operoni ya lac ya E. koli. Kwa kazi hii, walishinda Tuzo ya Nobel katika Physiolojia au Tiba mwaka wa 1965. Ingawa jeni za eukaryotiki hazipatikani kuwa operoni, operoni za prokaryotiki ni mifano bora ya kujifunza kuhusu udhibiti wa jeni kwa ujumla. Kuna baadhi ya makundi ya jeni katika eukaryotes ambayo hufanya kazi sawa na operoni. Kanuni nyingi zinaweza kutumika kwa mifumo ya eukaryotiki na kuchangia ufahamu wetu wa mabadiliko katika usemi wa jeni katika eukaryotes ambayo yanaweza kusababisha mabadiliko ya kiafya kama vile kansa.
Kila operon inajumuisha utaratibu wa DNA unaoathiri transcription yake mwenyewe; hizi ziko katika kanda inayoitwa kanda ya udhibiti. Eneo la udhibiti linajumuisha mtangazaji na kanda inayozunguka mtangazaji, ambayo sababu za transcription, protini zilizowekwa na jeni za udhibiti, zinaweza kumfunga. Sababu za transcription huathiri kisheria kwa polymerase ya RNA kwa promoter na kuruhusu maendeleo yake ya kuandika jeni za kimuundo. repressor - transcription sababu kwamba suppresses transcription ya jeni katika kukabiliana na kichocheo nje kwa kumfunga kwa mlolongo wa DNA ndani ya eneo udhibiti aitwaye operator, ambayo iko kati ya RNA polymerase kisheria tovuti ya promota na transcriptional kuanza tovuti ya kwanza miundo gene. Kufungia kisheria kimwili huzuia RNA polymerase kutoka kwa kuandika jeni za kimuundo. Kinyume chake, activator ni sababu ya transcription ambayo huongeza transcription ya jeni katika kukabiliana na kichocheo cha nje kwa kuwezesha RNA polymerase kisheria kwa promoter. Inducer, aina ya tatu ya molekuli ya udhibiti, ni molekuli ndogo ambayo inaamsha au huzuia transcription kwa kuingiliana na repressor au activator.
Katika prokaryotes, kuna mifano ya operons ambao bidhaa za jeni zinahitajika badala ya mara kwa mara na ambao maneno yake, kwa hiyo, hayawezi kudhibitiwa. Operoni hizo zinaonyeshwa kwa kikao, maana yake ni zimeandikwa na kutafsiriwa kwa kuendelea kutoa kiini na viwango vya kati vya mara kwa mara vya bidhaa za protini. Jeni hizo zinajumuisha enzymes zinazohusika katika kazi za utunzaji wa nyumba zinazohitajika kwa ajili ya matengenezo ya seli, ikiwa ni pamoja na replication ya DNA, kutengeneza, na kujieleza, pamoja na enzymes Kwa upande mwingine, kuna operons nyingine za prokaryotic ambazo zinaelezwa tu wakati zinahitajika na zinasimamiwa na waandamizaji, watendaji, na inducers.
Zoezi\(\PageIndex{1}\)
- Ni sehemu gani katika mlolongo wa DNA wa operon?
- Ni aina gani za molekuli za udhibiti zilizopo?
Udhibiti na Ukandamizaji
Operons Prokaryotic kawaida kudhibitiwa na kisheria ya repressors kwa mikoa operator, na hivyo kuzuia transcription ya jeni miundo. Watu kama hao huwekwa kama watu wanaoweza kukandamiza au watu wasioweza kuambukizwa. Operons repressible, kama tryptophan (trp) operon, kwa kawaida huwa na jeni encoding Enzymes zinazohitajika kwa njia biosynthetic. Kwa muda mrefu kama bidhaa ya njia, kama tryptophan, inaendelea kuhitajika na kiini, operon ya repressible itaendelea kuonyeshwa. Hata hivyo, wakati bidhaa za njia ya biosynthetic huanza kujilimbikiza kwenye seli, kuondoa haja ya kiini kuendelea kufanya zaidi, usemi wa operon unakandamizwa. Kinyume chake, operons inducible, kama operon ukosefu wa E. coli, mara nyingi huwa na jeni encoding enzymes katika njia kushiriki katika kimetaboliki ya substrate maalum kama lactose. Enzymes hizi zinahitajika tu wakati substrate kwamba inapatikana, hivyo kujieleza ya operons ni kawaida ikiwa tu mbele ya substrate.
Operesheni ya safari: Operesheni ya kukandamiza
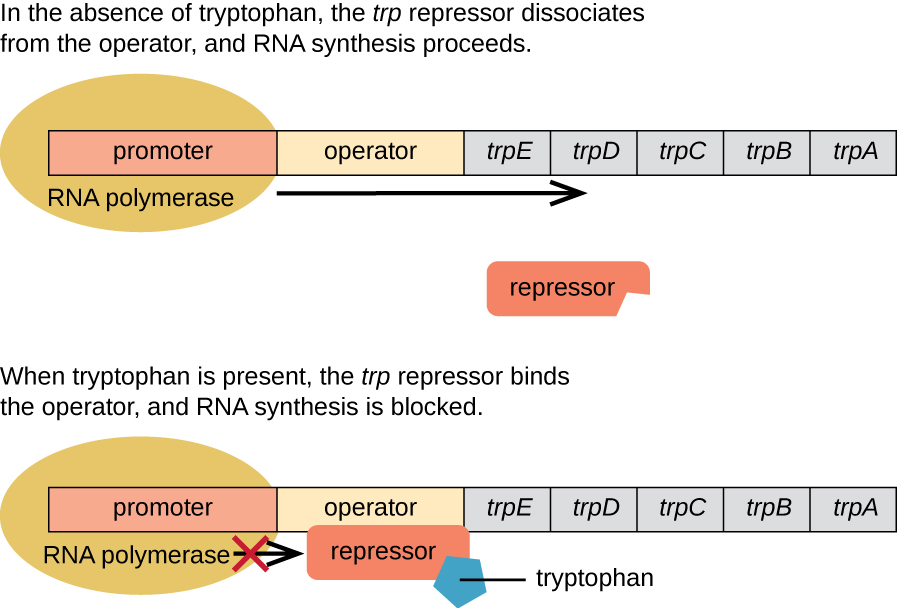
E. koli inaweza kuunganisha tryptophan kwa kutumia enzymes ambazo zimehifadhiwa na jeni tano za kimuundo ziko karibu na kila mmoja katika operon ya trp (Kielelezo\(\PageIndex{2}\)). Wakati tryptophan ya mazingira iko chini, operon imegeuka. Hii inamaanisha kuwa transcription imeanzishwa, jeni zinaelezwa, na tryptophan inatengenezwa. Hata hivyo, ikiwa tryptophan iko katika mazingira, operesheni ya safari imezimwa. Transcription haina kutokea na tryptophan si synthesized.
Wakati tryptophan haipo katika kiini, mkandamizaji yenyewe hakumfunga kwa operator; kwa hiyo, operon inafanya kazi na tryptophan inatengenezwa. Hata hivyo, wakati tryptophan hujilimbikiza katika seli, molekuli mbili za tryptophan hufunga kwenye molekuli ya trp ya kukandamiza, ambayo hubadilisha sura yake, na kuruhusu kuifunga kwa operator wa trp. Ufungaji huu wa fomu ya kazi ya mtetezi wa trp kwa operator huzuia polymerase ya RNA kutoka kwa kuandika jeni za kimuundo, kuacha kujieleza kwa operon. Hivyo, bidhaa halisi ya njia ya biosynthetic inayodhibitiwa na operon inasimamia usemi wa operon.
Tazama video hii ili ujifunze zaidi kuhusu operon ya safari.
Operesheni ya kukosa: Operesheni ya Inducible
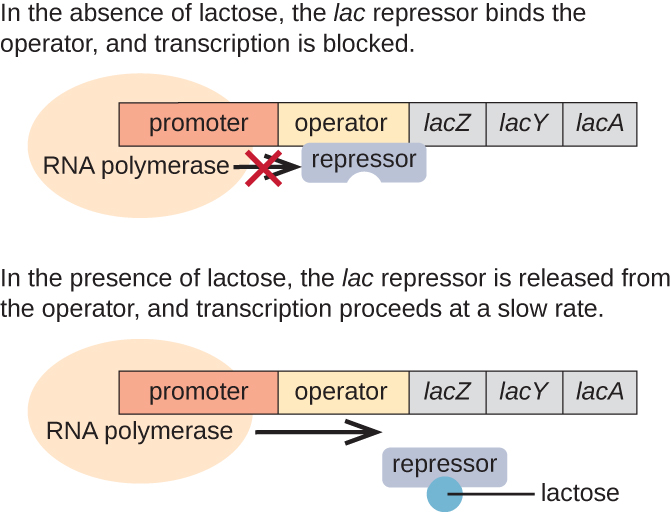
Operesheni ya kukosa ni mfano wa mtu anayeweza kuambukizwa ambayo pia inakabiliwa na uanzishaji kwa kutokuwepo kwa glucose (Kielelezo\(\PageIndex{3}\)). Operon ya lac inajumuisha jeni tatu za kimuundo zinazohitajika kupata na kutengeneza lactose ya disaccharide kutoka kwa mazingira, kuivunja ndani ya sukari rahisi glucose na galactose. Kwa mtu wa kukosa kuonyeshwa, lactose lazima iwepo. Hii mantiki kwa kiini kwa sababu itakuwa energetically kupoteza kujenga Enzymes mchakato lactose kama lactose haikupatikana.
Kutokana na kukosekana kwa lactose, kukandamiza lac ni wajibu wa eneo operator wa operon lac, kimwili kuzuia RNA polymerase kutoka transcribing jeni miundo. Hata hivyo, wakati lactose iko, lactose ndani ya seli inabadilishwa kuwa allolactose. Allolactose hutumika kama molekuli ya inducer, inayofunga kwa mkandamizaji na kubadilisha sura yake ili iweze tena kumfunga kwa DNA ya operator. Uondoaji wa mkandamizaji mbele ya lactose inaruhusu RNA polymerase kuhamia kanda ya operator na kuanza transcription ya jeni lac miundo.
Operon kukosa: Activation na Catabolite Activator Protini
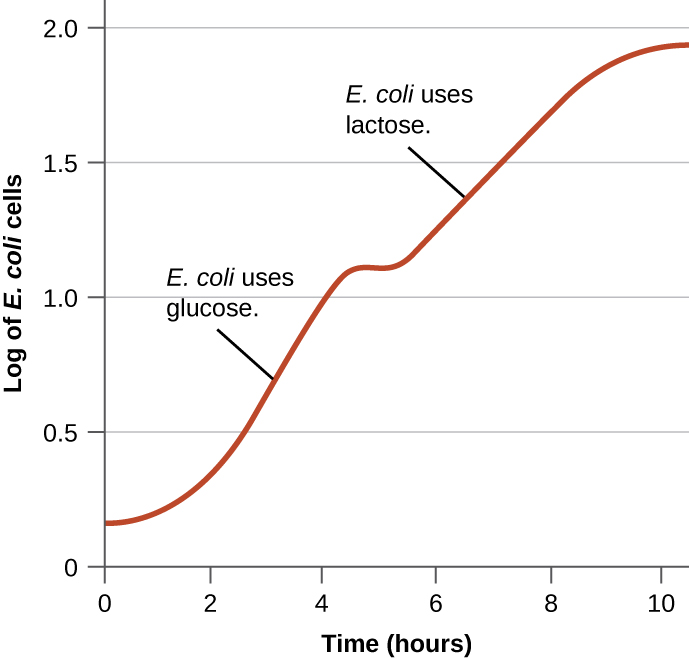
Bakteria huwa na uwezo wa kutumia substrates mbalimbali kama vyanzo vya kaboni. Hata hivyo, kwa sababu glucose kawaida hupendekezwa kwa substrates nyingine, bakteria zina taratibu za kuhakikisha kuwa substrates mbadala hutumiwa tu wakati glucose imepungua. Zaidi ya hayo, bakteria zina taratibu za kuhakikisha kwamba encoding enzymes za jeni kwa kutumia substrates mbadala zinaonyeshwa tu wakati substrate mbadala inapatikana. Katika miaka ya 1940, Jacques Monod alikuwa wa kwanza kuonyesha upendeleo kwa substrates fulani juu ya wengine kupitia masomo yake ya ukuaji wa E. coli wakati alipopandwa mbele ya substrates mbili tofauti wakati huo huo. Masomo hayo yanayotokana curves diauxic ukuaji, kama moja inavyoonekana katika Kielelezo\(\PageIndex{4}\). Ingawa glucose iliyopendekezwa ya substrate hutumiwa kwanza, E. coli inakua haraka na enzymes za kimetaboliki ya lactose hazipo. Hata hivyo, mara moja viwango vya glucose vimeharibika, viwango vya ukuaji hupungua, na kusababisha usemi wa enzymes zinazohitajika kwa kimetaboliki ya substrate ya pili, lactose. Angalia jinsi kiwango cha ukuaji katika lactose ni polepole, kama ilivyoonyeshwa na mwinuko wa chini wa curve ya ukuaji.
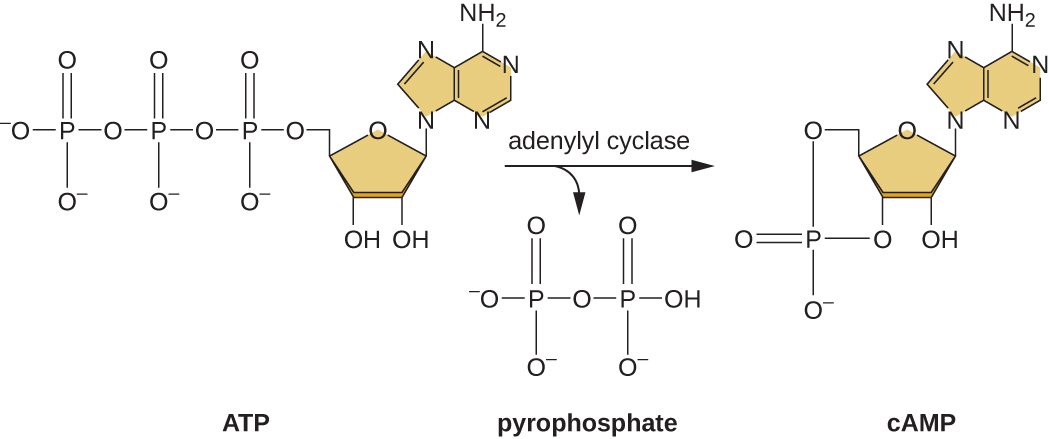
Uwezo wa kubadili kutoka kwa matumizi ya glucose hadi kwenye substrate nyingine kama lactose ni matokeo ya shughuli za enzyme inayoitwa Enzyme IIA (EIIA). Wakati viwango vya glucose kushuka, seli kuzalisha chini ATP kutoka catabolism (tazama Catabolism ya Wanga), na EIIA inakuwa phosphorylated. Fosforasi EIIA activates adenyll cyclase, enzyme kwamba waongofu baadhi ya ATP iliyobaki kwa mzunguko AMP (cAMP), mzunguko derivative ya AMP na muhimu ishara molekuli kushiriki katika glucose na nishati kimetaboliki katika E. coli. Matokeo yake, ngazi za cAMP zinaanza kuongezeka katika kiini (Kielelezo\(\PageIndex{5}\)).
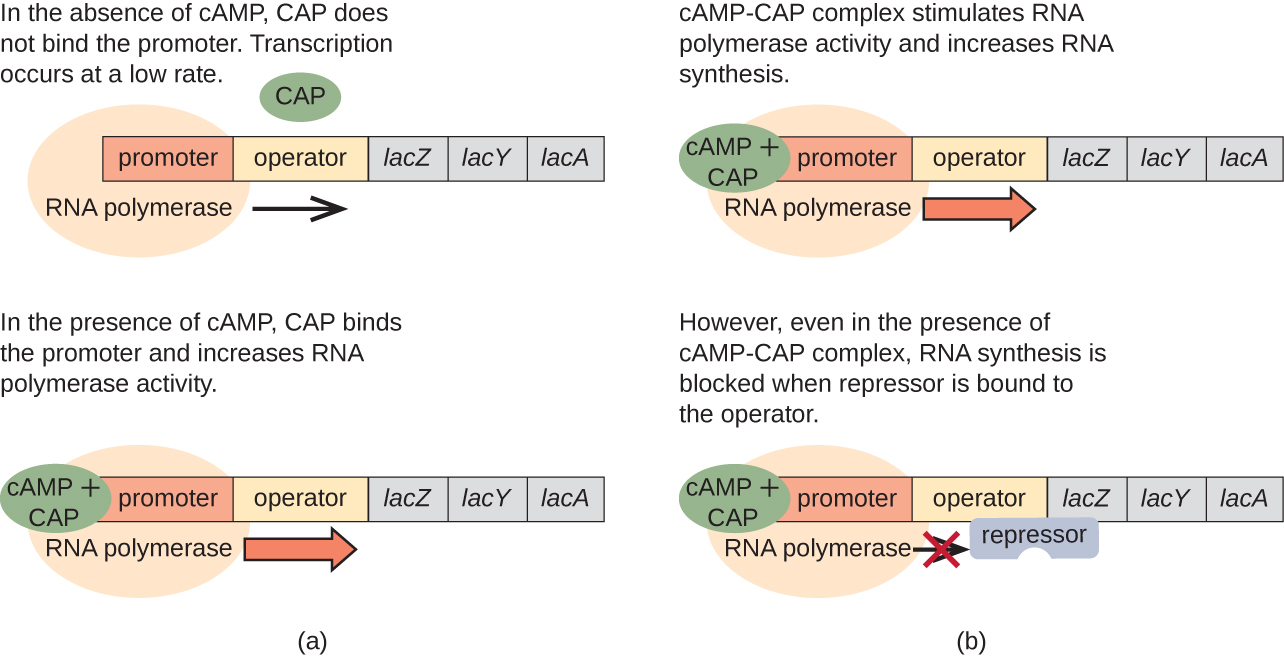
Operon lace pia ina jukumu katika kubadili hii kutoka kwa kutumia glucose kutumia lactose. Wakati glucose ni chache, kujilimbikiza CAMP unasababishwa na kuongezeka kwa shughuli adenylyl cyclase kumfunga kwa catabolite activator protini (CAP), pia inajulikana kama CAMP receptor protini (CRP). Ngumu hufunga kwa mkoa wa promoter wa operesheni ya lace (Kielelezo\(\PageIndex{6}\)). Katika mikoa ya udhibiti wa operons hizi, tovuti ya kisheria ya CAP iko juu ya tovuti ya kisheria ya RNA polymerase katika mtangazaji. Kufungwa kwa tata ya Cap-camp kwenye tovuti hii huongeza uwezo wa kumfunga wa RNA polymerase kwa mkoa wa promoter ili kuanzisha transcription ya jeni za kimuundo. Kwa hiyo, katika kesi ya operon ya lac, kwa transcription kutokea, lactose lazima iwepo (kuondoa protini ya kukandamiza lac) na viwango vya glucose lazima zimeharibika (kuruhusu kumfunga protini inayoamsha). Wakati viwango vya glucose ni vya juu, kuna ukandamizaji wa catabolite wa enzymes ya enzymes ya enzymes kwa metaboli ya substrates Kwa sababu ya viwango vya chini vya CAMP chini ya hali hizi, kuna kiasi cha kutosha cha tata ya Cap-camp ili kuamsha transcription ya operons hizi. Angalia Jedwali\(\PageIndex{1}\) kwa muhtasari wa udhibiti wa operon lac.
| Glucose | CAP hufunga | Lactose | Mkandamizaji hufunga | Transcription |
|---|---|---|---|---|
| + | — | — | + | Hapana |
| + | — | + | — | Baadhi |
| — | + | — | + | Hapana |
| — | + | + | — | Ndio |
Tazama mafunzo ya uhuishaji kuhusu kazi za lac operon hapa.
Zoezi\(\PageIndex{2}\)
- Ni nini kinachoathiri kisheria kwa mtetezi wa safari kwa operator?
- Je! Tabia ya protini ya kukandamiza lac imebadilikaje na lini?
- Mbali na kuwa repressible, ni kingine gani operon ukosefu umewekwa?
Majibu ya Kimataifa ya Prokaryotes
Katika prokaryotes, pia kuna viwango kadhaa vya juu vya udhibiti wa jeni ambavyo vina uwezo wa kudhibiti transcription ya operoni nyingi zinazohusiana wakati huo huo katika kukabiliana na ishara ya mazingira. Kikundi cha watu wote kudhibitiwa wakati huo huo huitwa regulon.
Alarmones
Wakati wa kuhisi shida inayotarajiwa, prokaryotes hubadilisha usemi wa aina mbalimbali za operons kujibu kwa uratibu. Wanafanya hivyo kwa njia ya uzalishaji wa alarmones, ambazo ni ndogo za nucleotide za intracellular. Alarmones hubadilisha ambayo jeni huelezwa na kuchochea usemi wa jeni maalum za kukabiliana na dhiki. Matumizi ya alarmones kubadilisha usemi wa jeni katika kukabiliana na dhiki inaonekana kuwa muhimu katika bakteria ya pathogenic. Katika kukutana na utaratibu wa ulinzi wa jeshi na hali nyingine kali wakati wa maambukizi, watu wengi encoding jeni virulence ni upregulated katika kukabiliana na ishara alarmone. Maarifa ya majibu haya ni muhimu kwa kuwa na uwezo wa kuelewa kikamilifu mchakato wa maambukizi ya vimelea vingi na kwa maendeleo ya matibabu ili kukabiliana na mchakato huu.
Mambo mbadala σ
Kwa kuwa subunit σ ya polymerase ya RNA ya bakteria inatoa maalum kuhusu wapromota ambao wanapaswa kuandikwa, kubadilisha sababu σ kutumika ni njia nyingine ya bakteria kwa haraka na kimataifa kubadilisha nini regulons ni transcribed kwa wakati fulani. Kipengele cha σ kinatambua utaratibu ndani ya promota ya bakteria, hivyo mambo tofauti ya σ kila mmoja atambua utaratibu tofauti wa promoter. Kwa njia hii, wakati kiini huhisi mazingira maalum ya mazingira, inaweza kujibu kwa kubadilisha ni kipengele gani σ kinachoonyesha, kudhalilisha zamani na kuzalisha moja mpya ya kuandika operons encoding jeni ambazo bidhaa zitakuwa na manufaa chini ya hali mpya ya mazingira. Kwa mfano, katika sporulating bakteria ya genera Bacillus na Clostridium (ambayo ni pamoja na vimelea vingi), kundi la mambo σ hudhibiti usemi wa jeni nyingi zinazohitajika kwa sporulation katika kukabiliana na ishara za kuchochea sporulation.
Zoezi\(\PageIndex{3}\)
- Je! Jina lililopewa mkusanyiko wa operons ambazo zinaweza kudhibitiwa kama kikundi?
- Ni aina gani ya kichocheo ingeweza kusababisha transcription ya tofauti σ sababu?
Mbinu za ziada za Udhibiti katika Bakteria: Kuzuia na Riboswitches
Ingawa wengi kujieleza jeni ni umewekwa katika ngazi ya transcription uanzishwaji katika prokaryotes, kuna pia taratibu za kudhibiti wote kukamilika kwa transcription pamoja na tafsiri wakati huo huo. Tangu ugunduzi wao, taratibu hizi zimeonyeshwa kudhibiti kukamilika kwa transcription na tafsiri ya operons nyingi za prokaryotic. Kwa sababu taratibu hizi zinaunganisha udhibiti wa transcription na tafsiri moja kwa moja, ni maalum kwa prokaryotes, kwa sababu taratibu hizi zinajitenga kimwili katika eukaryotes.
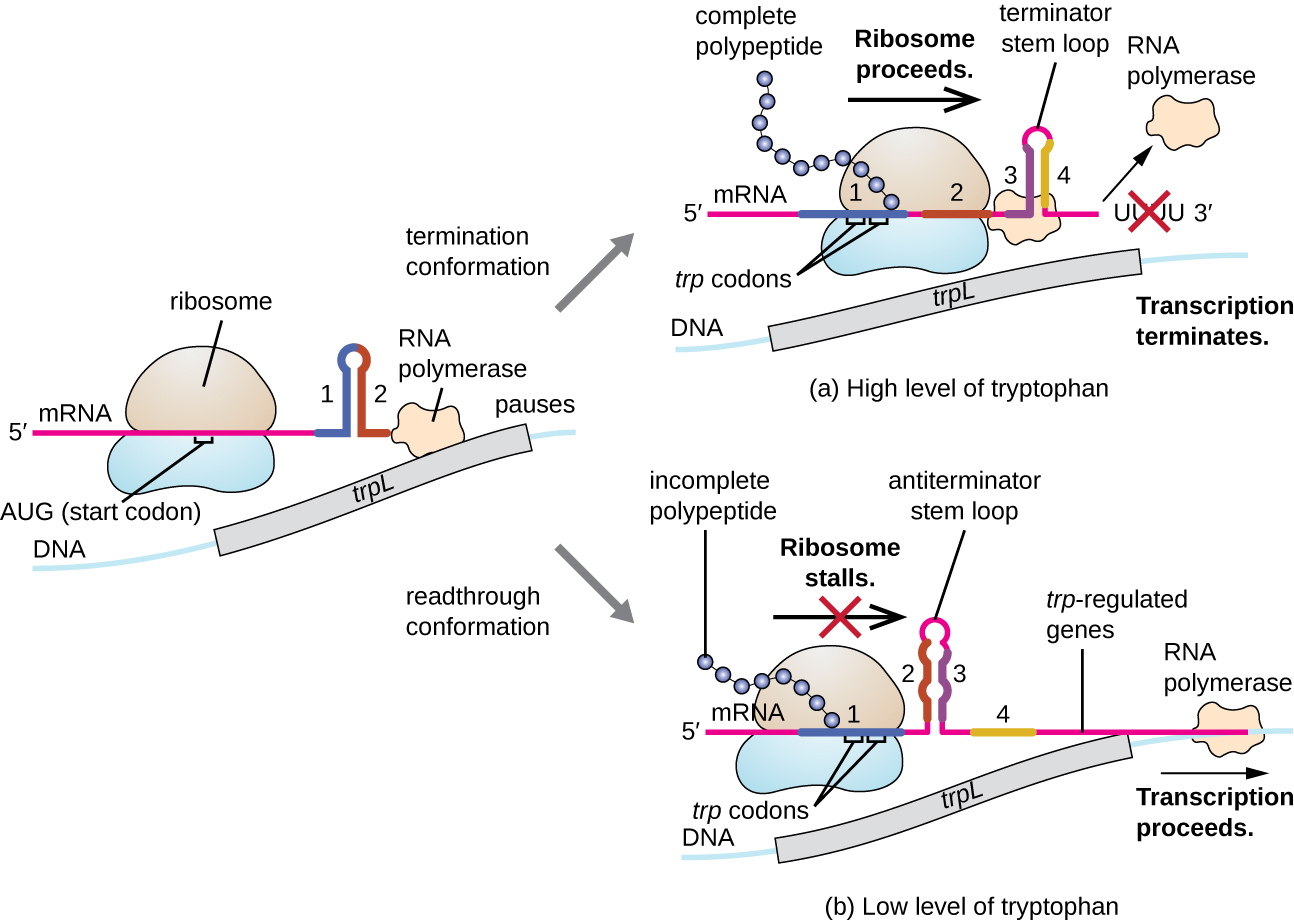
Moja kama mfumo wa udhibiti ni attenuation, ambapo sekondari shina kitanzi miundo sumu ndani ya 5' mwisho wa mRNA kuwa transcribed kuamua kama transcription kukamilisha awali ya mRNA hii itatokea na kama mRNA hii itatumika kwa ajili ya tafsiri. Zaidi ya utaratibu wa ukandamizaji wa transcriptional tayari kujadiliwa, attenuation pia hudhibiti usemi wa operon trp katika E. coli (Kielelezo\(\PageIndex{7}\)). Eneo la udhibiti wa trp operon lina mlolongo wa kiongozi unaoitwa TrPL kati ya operator na jeni la kwanza la kimuundo, ambalo lina stretches nne za RNA ambazo zinaweza jozi ya msingi na kila mmoja katika mchanganyiko tofauti. Wakati aina ya shina la kitanzi cha mwisho, transcription imekoma, ikitoa RNA polymerase kutoka mRNA. Hata hivyo, wakati aina antiterminator shina kitanzi, hii inazuia malezi ya terminator shina kitanzi, hivyo RNA polymerase inaweza transcribe jeni miundo.
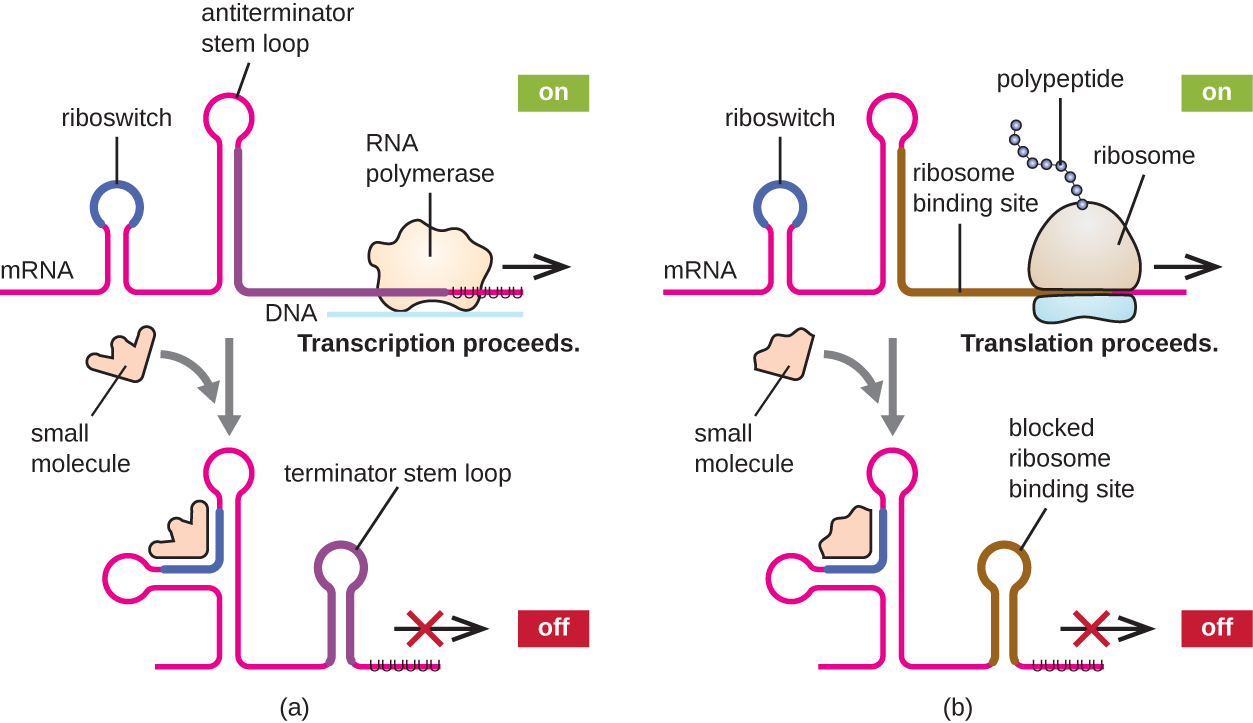
Utaratibu unaohusiana wa udhibiti wa wakati mmoja wa transcription na tafsiri katika prokaryotes ni matumizi ya riboswitch, kanda ndogo ya RNA isiyo ya coding inayopatikana ndani ya mwisho wa 5' wa molekuli fulani za mRNA za prokaryotic (Kielelezo\(\PageIndex{8}\)). Riboswitch inaweza kumfunga kwa molekuli ndogo ya intracellular ili kuimarisha miundo fulani ya sekondari ya molekuli ya mRNA. Kufungwa kwa molekuli ndogo huamua aina gani za muundo wa shina, na hivyo kushawishi kukamilika kwa awali ya mRNA na awali ya protini.
Mambo mengine yanayoathiri Ufafanuzi wa Gene katika Prokaryotes na Eukaryotes
Ingawa lengo la majadiliano yetu ya kudhibiti transcriptional kutumika operons prokaryotic kama mifano, eukaryotic transcriptional kudhibiti ni sawa kwa njia nyingi. Kama ilivyo katika prokaryotes, transcription ya eukaryotic inaweza kudhibitiwa kwa njia ya kumfunga mambo ya transcription ikiwa ni pamoja na repressors na activators. Jambo la kushangaza, eukaryotic transcription inaweza kuathiriwa na kisheria ya protini kwa mikoa ya DNA, aitwaye enhancers, badala mbali na jeni, kwa njia ya DNA looping kuwezeshwa kati enhancer na promota (Kielelezo\(\PageIndex{9}\)). Kwa ujumla, kusimamia transcription ni njia yenye ufanisi sana ya kudhibiti usemi wa jeni katika prokaryotes zote na eukaryotes. Hata hivyo, udhibiti wa usemi wa jeni katika eukaryotes katika kukabiliana na matatizo ya mazingira na seli unaweza kukamilika kwa njia za ziada bila kumfunga mambo ya transcription kwa mikoa ya udhibiti.
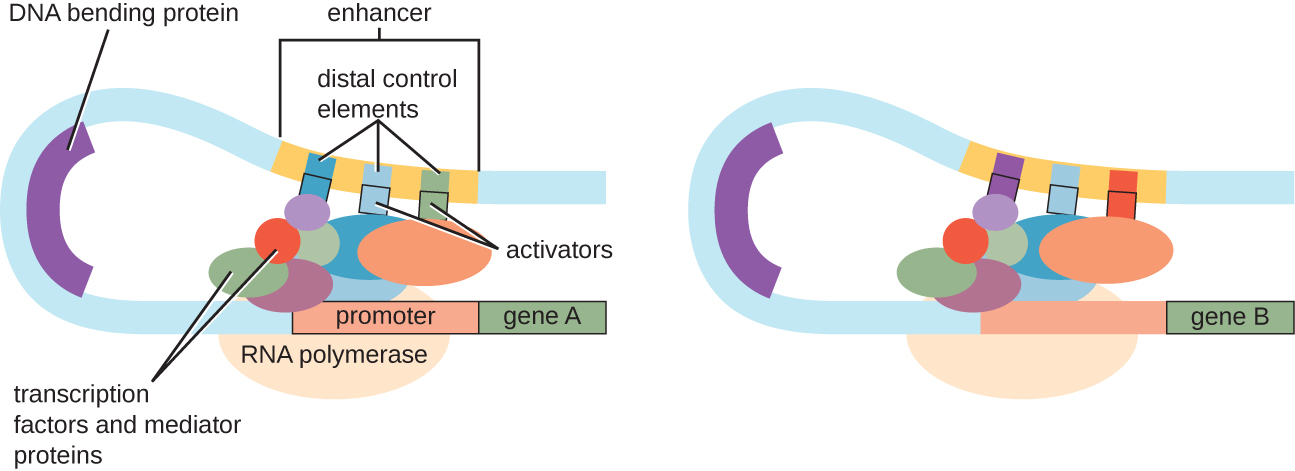
Udhibiti wa ngazi ya DNA
Katika eukaryotes, molekuli za DNA au histones zinazohusiana zinaweza kubadilishwa kwa kemikali kwa njia ya kuathiri transcription; hii inaitwa kanuni ya epigenetic. Methylation ya nucleotidi fulani ya cytosini katika DNA katika kukabiliana na mambo ya mazingira umeonyeshwa kuathiri matumizi ya DNA hiyo kwa transcription, huku methylation ya DNA kwa kawaida inahusiana na viwango vya chini vya kujieleza jeni. Zaidi ya hayo, katika kukabiliana na mambo ya mazingira, histone protini kwa ajili ya ufungaji DNA pia inaweza kemikali kubadilishwa kwa njia nyingi, ikiwa ni pamoja na acetylation na deacetylation, kushawishi hali ya ufungaji wa DNA na hivyo kuathiri upatikanaji wa huru jeraha DNA kwa transcription. Marekebisho haya kemikali wakati mwingine inaweza kudumishwa kwa njia ya raundi mbalimbali ya mgawanyiko kiini, na kufanya angalau baadhi ya mabadiliko haya epigenetic heritable.
Video hii inaeleza jinsi kanuni za epigenetic zinazodhibiti kujieleza kwa jeni.
Zoezi\(\PageIndex{}\)
- Ni nini kinachoacha au inaruhusu transcription kuendelea wakati uzuiaji unafanya kazi?
- Nini huamua hali ya riboswitch?
- Eleza kazi ya enhancer.
- Eleza njia mbili za kanuni za epigenetic katika eukaryotes.
Mtazamo wa Hospitali
Ingawa Mark alinusurika bout yake na fasciitis necrotizing, yeye sasa ingekuwa na upasuaji wa ngozi grafting, ikifuatiwa na tiba ya muda mrefu ya kimwili. Kulingana na kiasi cha misuli ya misuli aliyopoteza, haiwezekani kwamba mguu wake utarudi kwa nguvu kamili, lakini mtaalamu wake wa kimwili ana matumaini kwamba atapata tena matumizi ya mguu wake.
Upimaji wa maabara ulifunua wakala wa causative wa maambukizi ya Mark ilikuwa aina ya streptococcus ya kundi A (Kundi A strep). Kama inavyotakiwa na sheria, kesi ya Mark iliripotiwa kwa idara ya afya ya serikali na hatimaye kwa Vituo vya Kudhibiti na Kuzuia Magonjwa (CDC). Katika CDC, aina ya strep ya kundi A iliyotengwa na Mark ilichambuliwa vizuri zaidi kwa upinzani wa methicillin.
Methicillin upinzani ni jeni encoded na ni kuwa zaidi ya kawaida katika kundi A strep kupitia usawa uhamisho jeni. Katika fasciitis necrotizing, mtiririko wa damu kwa eneo la kuambukizwa ni kawaida mdogo kwa sababu ya hatua ya sumu mbalimbali encoded bakteria. Hii ni kwa nini kuna kawaida kidogo na hakuna kutokwa na damu kutokana na mtihani incision. Kwa bahati mbaya, sumu hizi za bakteria hupunguza ufanisi wa antibiotics ya ndani katika kusafisha maambukizi kutoka kwa ngozi na tishu za msingi, maana yake ni kwamba upinzani wa antibiotiki pekee hauelezei ufanisi wa matibabu ya Marko. Hata hivyo, tiba ya antibiotic ya intravenous ilitakiwa kusaidia kupunguza matokeo iwezekanavyo ya sepsis, ambayo ni matokeo ya kawaida ya fasciitis ya necrotizing. Kupitia uchambuzi wa jenomu na CDC ya aina iliyotengwa na Mark, kadhaa ya jeni muhimu za virulence zilionyeshwa kuwa encoded juu ya unabii, kuonyesha kwamba transduction ni muhimu katika uhamisho wa jeni usawa wa jeni hizi kutoka seli moja ya bakteria hadi nyingine.
Dhana muhimu na Muhtasari
- Gene kujieleza ni mchakato tightly umewekwa.
- Ufafanuzi wa jeni katika prokaryotes umewekwa kwa kiasi kikubwa katika hatua ya transcription. Ufafanuzi wa jeni katika eukaryotes unaongezewa zaidi baada ya transcriptionally.
- Prokaryotic jeni miundo ya kazi kuhusiana ni mara nyingi kupangwa katika operons, wote kudhibitiwa na transcription kutoka promota moja. Eneo la udhibiti wa operon linajumuisha promoter yenyewe na kanda inayozunguka promoter ambayo mambo ya transcription yanaweza kumfunga kushawishi transcription.
- Ingawa baadhi ya operons ni contitively walionyesha, wengi ni chini ya kanuni kupitia matumizi ya mambo transcription (repressors na activators). Mkandamizaji hufunga kwa operator, mlolongo wa DNA ndani ya eneo la udhibiti kati ya tovuti ya kisheria ya RNA ya polymerase katika mtangazaji na jeni la kwanza la miundo, na hivyo kuzuia kimwili transcription ya operons hizi. Activator hufunga ndani ya eneo la udhibiti wa operon, kusaidia RNA polymerase kumfunga kwa promoter, na hivyo kuimarisha transcription ya operon hii. Inducer inathiri transcription kupitia kuingiliana na repressor au activator.
- Mtu wa safari ni mfano wa classic wa operon repressible. Wakati tryptophan inakusanya, tryptophan hufunga kwa mkandamizaji, ambayo hufunga kwa operator, kuzuia transcription zaidi.
- Mtu wa lace ni mfano wa classic na mtu inducible. Wakati lactose iko kwenye seli, inabadilishwa kuwa allolactose. Allolactose hufanya kama inducer, kumfunga kwa mkandamizaji na kuzuia mkandamizaji kutoka kumfunga kwa operator. Hii inaruhusu transcription ya jeni miundo.
- Mtu wa kukosa pia ni chini ya uanzishaji. Wakati viwango vya glucose vimeharibika, baadhi ya ATP za mkononi hubadilishwa kuwa cAMP, ambayo hufunga kwa protini ya activator ya catabolite (CAP). Ngumu ya cap ya kambi hufanya transcription ya operator lace. Wakati viwango vya glucose ni vya juu, uwepo wake huzuia transcription ya operon kukosa na watu wengine kwa ukandamizaji wa catabolite.
- Molekuli ndogo za intracellular zinazoitwa alarmones zinafanywa kwa kukabiliana na matatizo mbalimbali ya mazingira, kuruhusu bakteria kudhibiti transcription ya kundi la operoni, inayoitwa regulon.
- Bakteria wana uwezo wa kubadili ni kipi σ cha polimerasi ya RNA wanayotumia katika kukabiliana na hali ya mazingira ili kubadilisha haraka na kimataifa ambayo regulons ni transcribed.
- Prokaryotes zina taratibu za udhibiti, ikiwa ni pamoja na uzuiaji na matumizi ya riboswitches, ili kudhibiti wakati huo huo kukamilika kwa transcription na tafsiri kutoka kwa nakala hiyo. Njia hizi zinafanya kazi kwa njia ya kuundwa kwa loops za shina katika mwisho wa 5' wa molekuli ya mRNA inayounganishwa sasa.
- Kuna pointi za ziada za udhibiti wa kujieleza jeni katika prokaryotes na eukaryotes. Katika eukaryotes, udhibiti wa epigenetic na mabadiliko ya kemikali ya DNA au histones, na udhibiti wa usindikaji wa RNA ni njia mbili.